

Bioprinting organoids for functional cardiac constructs: Progress and unmet challenges

Developing physiologically relevant cardiac-engineered in vitro models has been a longstanding challenge in cardiac tissue engineering. Bioprinting technologies have been utilized to recreate the complex architecture of the human heart via the precise placement of cells and biomaterials. Concurrently, self-organizing cardiac organoids have emerged as powerful tools for developing cardiac tissues accurately mimicking the heart’s biological composition. This review explores the merging of these two rapidly evolving fields to produce functionally mature engineered cardiac tissues. Together, bioprinting can provide spatial control and mechanical support to guide cardiac self-organization, including strategies to directly print cardioids or incorporate them as modular units, while cardioid differentiation protocols promote multi-cellular complexity and developmental relevance to improve the functionality of engineered cardiac constructs. In this review, we discuss the key processing challenges and goals across the bioprinting workflow—spanning pre-processing, processing, and post-processing—and evaluate how they intersect with cell viability, structural integrity, and electromechanical function. We then explore the formation and functional features of self-organized cardioids, outlining major differentiation protocols, signaling cues, and functional outcomes. Finally, we propose a convergence between bioprinting and cardioid technologies to produce the next generation of in vitro cardiac models.
1. Introduction
As the leading cause of death globally, cardiovascular disease (CVD) encompasses a range of disorders affecting the heart and blood vessels, including coronary heart disease, valvular heart disease, and congenital heart disease, claiming over 20 million lives annually.1 Over the past three decades, CVD-related deaths have increased by 60% and continue to rise.2 In the United States, more than half of the adult population experiences some form of CVD, ranging from early onset to late-stage disease.3 The heart’s minimal regenerative capacity makes it highly susceptible to irreversible damage, often leading to heart failure.4 Alarmingly, projections estimate that over 8 million people in the United States will suffer from heart failure by 2030.5 By 2050, the annual healthcare costs associated with CVD—currently $393 billion—are expected to rise to $1.49 trillion.6
As the CVD prevalence continues to rise, researchers have increasingly relied on advanced in vitro cardiac models as essential tools for developmental and disease modeling, drug testing, and regenerative medicine. Diverse cardiac tissue models—such as two-dimensional (2D) cultures, engineered three-dimensional (3D) constructs, and animal models—have been developed, but none fully replicate the structural and functional complexity of native heart tissues.7 The structural and biological complexity of the human heart presents significant challenges in engineering functional cardiac tissue.8 Technologies such as 3D bioprinting, an additive manufacturing process that deposits cells, biomaterials, and bioactive molecules in a spatially controlled manner, have been used to fabricate diverse cardiac constructs, ranging from simplified 2D myocardial sheets to complex 3D miniature hearts.9,10 Many bioprinting modalities have been explored in cardiac applications, each offering unique advantages depending on the intended application. However, regardless of modality, bioprinted cardiac constructs often fail to fully replicate native cardiac function due to limitations in cellular organization, vascularization, and functional maturation.7,9,10
Organoids are self-organizing 3D tissues derived from stem cells that recapitulate aspects of native tissue architecture and function. Cardiac organoids, sometimes referred to as cardioids, represent promising advancements in cardiac in vitro modeling.11 Through guided stem cell differentiation, cardioids, unlike pre-patterned tissue constructs, exhibit self-organizing properties, allowing them to form physiologically relevant cardiac structures, including chamber-like morphologies without predefined scaffolds.12 This developmentally driven self-organization makes cardioids valuable for studying cardiogenesis and congenital diseases. However, despite their advantages, cardioids remain limited in their applicability to adult cardiac applications due to microscale size, immaturity, poor vascularization, and lack of reproducibility.12–14
Merging bioprinting with cardioid technology could overcome the challenges of generating scalable, vascularized, and functionally mature cardiac tissues. By using bioprinting as a tool to guide cardiac structures into forming adult-like structures through self-organization, physiologically relevant and mature, engineered cardiac tissue can be generated in a scalable and highly reproducible manner. This review will first examine key aspects of the cardiac bioprinting workflow, including its use in fabricating myocardial constructs, heart valves, vascular grafts, etc. Next, we will provide a detailed discussion of cardioids, including their defining features, current applications, and limitations. Finally, we will assess recent progress toward bioprinting cardioids, outline the challenges involved, and propose potential solutions integrating bioprinting and cardioid technology.
2. Cardiac bioprinting
The bioprinting process consists of three key stages: pre-processing, processing, and post-processing (Figure 1). This section will explore critical considerations for each of these stages that are essential for obtaining prints with structural integrity and functional maturation, focusing on bioink properties, cell selection, bioprinting techniques, and methods for construct maturation.

Figure 1. Process map of cardiac tissue bioprinting. The bioprinting workflow consists of three primary stages: pre-processing, processing, and postprocessing. Pre-processing includes CAD selection/generation, cell selection, and bioink preparation, where biomaterials, bioactive molecules, and cells are combined. Processing involves the fabrication of cardiac constructs using various bioprinting modalities, including but not limited to droplet-based, extrusion-based, and light-based bioprinting. Post-processing focuses on tissue maturation through crosslinking, incubation, and maturation strategies such as bioreactor culturing. Created with BioRender.com. Abbreviation: CAD: Computer-aided design.
2.1. Pre-processing
Pre-processing involves all the preparatory steps necessary before the printing begins. First, the shape of the printed object is determined by computer-aided design (CAD) models, which can be manually designed or derived from medical imaging files, such as computed tomography or magnetic resonance imaging (MRI). Manually designed CAD models are generated directly within the software using basic modeling operations—such as sketching, extrusion, lofting, and cutting—to construct custom geometries without reliance on anatomical imaging sources. While manually designed CAD models are suitable for simple geometries such as strips, rings, and tubes, more anatomically complex models—such as valves, coronary arteries, and ventricles—are best generated from medical imaging data.15 Once an appropriate design is selected, a bioink that contains the necessary cellular and biomaterial components to support tissue function, can be selected.
2.1.1. Cells
The development, functionality, and maturation of cardiac tissue rely on a complex network of signaling and interactions among various cardiac cells.16,17 The major cardiac cell types—cardiomyocytes (CMs), endothelial cells (ECs), and cardiac fibroblasts (CFs)—comprise approximately 90% of the heart’s cellular population, while the remaining population includes vascular smooth muscle cells (VSMCs), immune cells, pericytes, adipocytes, and cardiac neurons.18 A summary of the major cardiac cells, their functions, and supporting cell types can be found in Figure 2.

Figure 2. Major and supporting cardiac cells. Cardiomyocytes are responsible for myocardial contractility and electrical conduction. Cardiac fibroblasts secrete ECMs and provide structural support. Endothelial cells line blood vessels, enabling vascularization and nutrient exchange. Vascular smooth muscle cells regulate vessel tone and allow arterial vasoconstriction/vasodilation. Supporting cell types, including pericytes, adipocytes, and macrophages, contribute to cardiac homeostasis and tissue remodeling. Created with BioRender.com. Abbreviation: EMC: Extracellular matrix.
For cardiac tissue engineering applications, pluripotent stem cells (PSCs), particularly induced pluripotent stem cells (iPSCs), are widely used due to their scalability and ability to differentiate into multiple cardiac lineages. While embryonic stem cells (ESCs) remain in use, iPSCs provide an ethical alternative, as they are derived and reprogrammed from somatic cells rather than embryonic tissue.19 During early cardiac engineering, CMs were once considered the only necessary cell type for functional cardiac tissue, resulting in efforts to prioritize optimizing PSC-CM differentiation and leading to a surge in protocols aimed at improving efficiency and reproducibility across multiple cell lines.20–22 Many bioprinted constructs have successfully generated contractile tissue using only iPSC/ ESC-derived CMs.23–32 Although CMs are critical for generating contraction and electrical signal propagation, CM interactions with non-myocyte populations are essential for functionality, as the absence of supporting cells compromises structural integrity, electromechanical function, and tissue longevity, and leads to pathological disruption. The sole focus of differentiation protocol development is no longer just on CMs; other cardiac cells—ECs in particular—are now receiving more attention.
Cardiac tissue is one of the most metabolically demanding tissues, requiring large amounts of oxygen and nutrient exchange to remain viable.33 To meet demands, cardiac tissue relies on dense vascular networks which are largely made up of ECs. Due to their availability and low cost, human umbilical vein endothelial cells (HUVECs) are commonly used in cardiac constructs. However, their venous phenotype limits their ability to form arterial vessels which are essential for effective vasculature.34–36 In contrast, PSC-derived ECs can differentiate into capillaries, arteries, and veins, making them essential for vascularization.37 In addition to ECs, VSMCs play a crucial role in regulating blood pressure and flow, maintaining vessel tone, and repairing injured arteries. PSC-EC and PSC-VSMC differentiation protocols are making swift progress in development, though not as abundant as CM protocols.38–41
While fibroblasts generally provide mechanical support and enable extracellular matrix (ECM) remodeling, CFs have specialized roles in heart tissue. They produce heart-specific ECM, secrete specific growth factors, and signaling molecules that promote CM alignment, maturation, and viability, regulate fibrosis to prevent excessive scarring, and are adapted to respond to cardiac-specific mechanical and electrical forces.42 Yet, many cardiac constructs continue to rely on primary CFs such as human dermal fibroblasts which lack the ability to perform the specialized tissue-specific tasks that CFs can.43–45 CF differentiation protocol development lags behind those of other major cardiac cells.46–48 Therefore, it is not uncommon to see bioink formulations using derived CMs and/or derived ECs with primary fibroblasts.49–54
Generally, using cardiac-derived cells is advantageous for situations where precise ratios of the major cardiac lineages are needed; however, completely recapitulating the native cardiac cellular composition remains challenging. Despite their relatively small populations, other cardiac cells, such as pericytes and macrophages, play critical roles in regulating major cell functions.55,56 However, differentiation protocols for these supporting cells remain limited, making their incorporation into a construct challenging. A potential solution involves differentiation protocols that simultaneously generate multiple cardiac lineages from early progenitors, like approaches commonly used in cardiac organoid development, which we will explore in later sections.
Beyond achieving the appropriate cellular composition, maintaining cardiac function during and after bioprinting requires recreating the biochemical and mechanical cues of the native environment. CM contractility and electrical propagation depend on alignment and gap junction formation, while ECs require signals that promote vessel formation and sustained metabolic activity. Fibroblasts and other stromal cells must also remain responsive to dynamic mechanical and biochemical inputs to preserve ECM structure and tissue organization. These functional attributes are supported not only by spatial patterning and biomaterial properties, but also by the incorporation of bioactive molecules that promote vascularization, matrix
remodeling, and CM maturation—factors that will be further explored in subsequent sections.
2.1.2. Bioink design
Unlike traditional 3D printing, which uses plastics or metals, bioprinting relies on bioinks—formulations that combine cells with biomaterials, and sometimes bioactive molecules, to fabricate 3D tissues. When designing a bioink, several key properties must first be considered to ensure successful fabrication and functional performance. Biocompatibility is essential for supporting cell viability, proliferation, and functionality without inducing cytotoxicity or immune responses. Printability determines the bioink’s ability to easily deposit while maintaining structural integrity after printing. In extrusion-based techniques, this requires shear-thinning properties, viscoelasticity, and controlled gelation kinetics to ensure smooth extrusion and post-printing architecture.57,58 In extrusion-based bioprinting, decreasing nozzle diameter improves resolution but increases pressure, which can reduce cell viability and lead to nozzle clogging. Optimizing bioink formulations—along with adjustments to printing parameters such as pressure, speed, and nozzle size—has been shown to enhance structural fidelity without minimizing shear stress-induced damage.59,60 For non-extrusion techniques, bioinks should have low viscosity to prevent nozzle clogging and improve deposition accuracy.
Mechanical properties must be optimized to mimic myocardial stiffness (8–17 kPa) to provide structural support and biomechanical cues essential for cardiac function.61 Additionally, bioinks must maintain adequate porosity to facilitate nutrient and oxygen diffusion throughout the construct. Biodegradability should be tuned to match the rate of tissue remodeling, ensuring that the scaffold provides support without premature degradation. Lastly, bioactivity is crucial for mimicking the cardiac ECM, by promoting cell attachment, proliferation, differentiation, and angiogenesis, all of which contribute to tissue function and maturation.57,59 Achieving these ideal bioink properties requires careful selection of appropriate biomaterials that balance biocompatibility, mechanical support, and biological signaling.
Hydrogels, which are 3D crosslinked polymer networks that retain water and mimic the ECM, are among the most widely used biomaterials for cardiac tissue engineering.62 These hydrogels can be composed of natural, synthetic, or hybrid polymers, each of which offers unique properties for cardiac tissue engineering. Natural biomaterials are derived from biological sources and are favored for their high biocompatibility and ability to mimic the ECM. Commonly used natural biomaterials for cardiac applications include alginate, decellularized extracellular matrix (dECM), and ECM components such as collagen, fibrin, gelatin, and Matrigel®, which are summarized in Table 1.
Table 1. Summary of biomaterials and bioactive molecules commonly used in cardiac bioprinting
| Biomaterial/biomolecule | Type | Key properties/functions | References |
|---|---|---|---|
| Alginate | Natural biomaterial | Biocompatible, easy gelation; used as a hydrogel scaffold to support cell encapsulation | 34,35,77 |
| Collagen | Natural biomaterial | Natural ECM component with RGD motifs; promotes cell attachment and tissue remodeling | 24,88 |
| dECM | Natural biomaterial | Retains native cardiac ECM composition with proteins and GAGs; recreates physiological microenvironment | 54 |
| Fibrin | Natural biomaterial | Pro-angiogenic, supports vascular network formation and myocardial regeneration | 34,53,67 |
| Gelatin | Natural biomaterial | ECM-mimicking; often modified to enhance printability and mechanical strength | 67 |
| GelMA | Natural biomaterial (modified) | Photocrosslinkable and bioactive; enables engineering of structured tissues | 23,25,32,35,51,89–91 |
| Matrigel® | Natural biomaterial | Rich in ECM proteins and growth factors; supports early cardiac tissue formation | 88 |
| PEG | Synthetic biomaterial | Highly tunable mechanical properties, bioinert polymer; used for creating customizable, hydrogel scaffolds | 34 |
| PEGDA | Synthetic biomaterial (modified) | PEG derivative with acrylate groups; allows photocrosslinkable hydrogel formation | 51,77 |
| VEGF | Bioactive molecule | Growth factor; stimulates EC proliferation and vascularization | * |
| FGF | Bioactive molecule | Growth factor; Promotes fibroblast and EC development for tissue remodeling | * |
| TGF-βi | Bioactive molecule | Modulate fibrosis and promote epicardial and fibroblast lineage specification | * |
*Used in media not bioink formulation
Abbreviations: dECM: Decellularized extracellular matrix; ECM: Extracellular matrix; FGF: Fibroblast growth factor; GAG: Glycosaminoglycan; GelMA: Gelatin methacryloyl; PEG: Polyethylene glycol; PEGDA: Polyethylene glycol diacrylate; RGD: Arginylglycylaspartic acid; TGF-βi: Transforming growth factor beta inhibitor; VEGF: Vascular endothelial growth factor.
Alginate is a naturally occurring polysaccharide that is highly biocompatible with a multitude of cells and other biomaterials. Alginate is used for a variety of biomedical applications due to its gelation properties and ease of modification.63 Derived from brown algae, alginate does not have naturally occurring cell-binding sites and must be modified with adhesive ligands to promote cell attachment.62 Alginate is often described as a blank slate biomaterial, making it particularly useful for maintaining the current cellular state. Collagen is the most abundant component of the ECM. As a bioink, collagen is commonly used due to its high biocompatibility, presence of cell-attachment-encouraging Arginylglycylaspartic acid (RGD) cell binding sites, enzymatic degradability, and temperature-dependent gelation.64 However, its low mechanical strength remains a significant limitation, especially for bioprinting.65 Fibrin, a protein involved in the coagulation cascade, has been utilized in bioprinting due to its applications in wound healing and its intrinsic angiogenic properties.66 Fibrin- based biomaterials facilitate the proliferation and spreading of multiple cardiac cells.67–69 Gelatin is a derivative of collagen that retains collagen’s biocompatibility, biodegradability, and ability to mimic the ECM.62 Like collagen, it is widely used in bioprinting despite its limited mechanical properties and is often modified to improve printability. Decellularized ECM-based bioinks are the most effective at recapitulating native tissues’ complex composition of proteins, glycosaminoglycans, proteoglycans, and other cardiac components.70 Decellularization techniques use chemical, physical, and/or biological agents to remove cells and immunogenic components, leaving behind a supportive scaffold for promoting cell adhesion, proliferation, migration, and differentiation.58 Matrigel® and other similar basement membranes such as Geltrex® and Cultrex® are widely utilized in both 2D and 3D cultures.71 They are highly compatible with cells due to their composition of ECM components such as laminin, collagen type IV, entactin, and heparan sulfate proteoglycans, and growth factors like transforming growth factor beta (TGF-β) and fibroblast growth factor (FGF). Batch variability, weak mechanical properties, and risk of inducing tumorigenicity limit Matrigel® bioink formulations.72,73
Chemically modifying natural biomaterials can improve certain properties. For example, the addition of methacrylate groups (–CH=CH2) can allow some biomaterials to become photocrosslinkable. Gelatin methacrylate (GelMA), also known as gelatin methacryloyl, is synthesized by reacting the primary amine (–NH2) and hydroxyl (–OH) groups of the gelatin with methacryloyl groups (–CO–CH=CH2) from methacrylic anhydride (C4H6O3). Like collagen, GelMA mimics the native ECM and promotes cell attachment, proliferation, and differentiation. However, GelMA offers superior printability due to its ability to be crosslinked by light and its tailorable mechanical properties.74 GelMA is often used in combination with other biomaterials to provide additional pro-regenerative stimulation through biological signals necessary for proliferation, differentiation, and other cell- to-cell interactions, and improve both printability and mechanical stability.23,35,70
Synthetic biomaterials offer several advantages in bioprinting. Their tunable chemical structure allows for precise control over degradation rates, mechanical properties, and crosslinking kinetics. They also exhibit excellent batch-to-batch consistency, long-term storage stability, and scalability for manufacturing applications. Furthermore, synthetic polymers can be readily functionalized with peptides, growth factors, or chemical groups to enhance cell adhesion and bioactivity. However, challenges including toxic byproducts, poor cell encapsulation, and a lack of natural adhesion sites limit their usage.75 Additionally, many synthetic polymers are inherently brittle and lack the elasticity required to replicate the biomechanical properties of the native myocardium.9 An exception to many of these limitations is polyethylene glycol (PEG), a polymer widely used in bioprinting and 3D culturing due to its hydrophilicity, tunability, and biocompatibility.75,76
Modified synthetic bioinks such as PEG hybrids are more widely used than pure synthetics in bioprinting. PEG-diacrylate (PEGDA), a PEG derivative commonly utilized for cardiac bioprinting, is capable of ultraviolet (UV) or visible light crosslinking due to the replacement of PEG’s hydroxyl (–OH) end groups with acrylate (–C=C–COO–) groups, enhancing structural stability.77,78 Cell adhesion-enhancing RGD peptides have been used to functionalize synthetic hydrogels and promote cardiac tissue formation.79,80 However, it is more common for researchers to employ hybrid bioink formulations of both natural and synthetic biomaterials to achieve bioinks with good printability and biocompatibility.
Beyond optimizing material composition alone, incorporating bioactive molecules into bioinks offers an additional strategy to guide cell behavior, promote differentiation, and enhance vascularization. While they are not always included in bioink formulations, bioactive molecules are critical for cardiac tissue engineering as they regulate cell attachment, proliferation, differentiation and maturation.
While early-stage differentiation factors such as CHIR99021(Chir)—a glycogen synthase kinase 3β inhibitor that triggers mesodermal induction of stem cells into progenitors—are difficult to incorporate due to concentration and timing sensitivities, later-stage growth factors such as vascular endothelial growth factor (VEGF) have been successfully encapsulated within hydrogels to support endothelialization and vascularization.38,39,81–83 Studies on controlled VEGF release from gels support the notion that VEGF is capable of supporting cellular proliferation and migration of encapsulated cells.84,85 These findings have contributed to the emergence of commercially marketed VEGF bioinks such as Sigma- Aldrich’s INVIVO-GEL VEGF and Amerigo Scientific’s Gel4Cell®-VEGF.
FGF and TGF-β inhibitors (TGF-βi), while not yet incorporated into commercial bioinks, have also been used to functionalize biomaterials and guide differentiation toward epicardial and fibroblast lineages.46–48,86,87 As the field progresses toward more heterogeneous constructs, controlled delivery of differentiation signals will likely become an essential component of next-generation cardiac bioinks.
2.2. Processing
To fabricate functional in vitro cardiac models, it is essential to balance structural precision, biomaterial selection and print fidelity. However, several processing challenges must be addressed to ensure the long-term viability, maturation, and functionality of engineered cardiac tissues. These include achieving physiological cell densities, maintaining mechanical stability, replicating anisotropic architectures, and integrating vascular networks (Figure 3).

Figure 3. Goals and challenges in engineering cardiac tissues. Engineering functional cardiac tissues require addressing several key challenges: achieving physiological cellular densities to enable proper electrical coupling and contraction; maintaining mechanical stability in macroscale constructs to preserve shape and function; replicating the anisotropic architecture of native myocardium to support directional signal propagation and coordinated contraction; and integrating vascular networks to supply oxygen and nutrients beyond the diffusion limit. Created with BioRender.com.
Bioprinting is broadly categorized into three modalities—extrusion-based bioprinting (EBB), light-based bioprinting (LBB), and droplet-based bioprinting (DBB), each offering distinct advantages in overcoming these challenges. These techniques have enabled the fabrication of a wide range of cardiac structures including patches, engineered muscle tissues, valves, chambers, and miniature whole-heart models.24,25,32,34,49–51,53,54,67,68,77,88–91
EBB continuously deposits high-viscosity bioinks using pneumatic pressure, pistons, or screw-driven forces, enabling high-cell-density, macroscale cardiac tissues. This technique is widely used for patches, engineered cardiac tissues, and other large-scale cardiac constructs, although it struggles with microarchitectural fidelity and microscale vascularization.92
LBB uses patterned light exposure to crosslink photoactive bioinks into 3D structures. Projection-based LBB techniques such as digital light processing (DLP) and volumetric bioprinting, enable high-resolution cardiac tissue fabrication and excel fabrication of microvasculature, patterned cellular and biomaterial alignment, and intricate cardiac architectures. However, they lack scalability and require specific bioink formulations that limit their versatility.93,94
DBB ejects discrete, cell-laden droplets via thermal, piezoelectric, or electromagnetic forces, making it ideal for high-precision cardiac cell patterning, endothelialization, and disease modeling. However, its reliance on low- viscosity bioinks limits structural stability, making it unsuitable for printing large or thick cardiac tissues.95
Table 2 summarizes the key characteristics of the primary bioprinting modalities, highlighting their deposition mechanisms, resolution, speed, bioink compatibility, and major advantages and limitations. This section explores how these distinctions within each modality address key structural, cellular and functional challenges in cardiac engineering.
Table 2. Cardiac bioprinting modalities
| Modality | Deposition mechanism | Resolution | Speed | Bioink compatibility | Advantages | Limitations |
|---|---|---|---|---|---|---|
| EBB | Continuous deposition of high-viscosity bioinks using pneumatic, piston, or screw-driven extrusion | 200–1000 µm | Slow–moderate | Wide range (natural, synthetic, hybrid) | High-cell-density printing, scalability, multimaterial printing | Limited resolution, poor microscale vascularization, shear stress-induced cell damage |
| LBB94,96 | Patterned light exposure to crosslink photoactive bioinks | 10–50 µm (DLP) < 10 µm (μCOP) | Fast | Requires photoactive bioresins | High resolution, patterned microvasculature, anisotropic tissue alignment | Limited scalability, bioink restrictions, cytotoxicity (UV light exposure) |
| DBB95 | Discrete bioink droplet ejection via thermal, piezoelectric, or electromagnetic forces | 10–50 µm | Fast | Low-viscosity bioinks | High precision for cell patterning, endothelialization | Low structural stability, unsuitable for bulk tissue fabrication |
Abbreviations: DBB: Droplet-based bioprinting; DLP: Digital light processing; EBB: Extrusion-based bioprinting; LBB: Light-based bioprinting; μCOP: Micro-continuous optical printing; UV: Ultraviolet.
2.2.1. Achieving physiological cellular density
The adult heart has an average cellular density of 30–40 million cells/mL.97 Direct cell–cell contact is a critical factor for CM differentiation and synchronized electrical propagation; therefore, matching these densities is critical to achieving functional constructs. Some studies have aimed to directly match bioink’s formulation to the heart’s cellular density prior to printing while others have relied on the proliferation of stem cells to reach confluency within the printed construct post-printing.23,88,98 Achieving these densities in constructs can pose a significant challenge. EBB is best suited for high-cell-density bioprinting, able to handle bioinks ranging from 106–108 cells/mL, whereas DBB can only handle around 106 cells/mL and LBB 106–107 cells/mL. Additionally, EBB can handle high- viscosity bioinks, including hydrogels which are optimal for cell encapsulation. However, printing at high density is not without tradeoffs. Both high biomaterial viscosity and high cell density increase the mechanical resistance of bioinks when extruded. The combination of the two significantly increases the amount of stress exerted on the cells. PSCs are particularly susceptible, often experiencing cell death, unintended differentiation, or loss of pluripotency.60 Mitigating shear stress-induced damage requires a balance of optimizing printing parameters such as extrusion pressure, nozzle diameter, and bioink viscosity. In contrast, LBB and DBB require low-viscosity bioinks to prevent light-scattering issues in LBB and nozzle clogging in DBB.98 As a result, LBB and DBB printers tend to report higher cell viability than EBB methods.
2.2.2. Maintaining mechanical stability in macroscale prints
Cardiac tissue engineering has yet to achieve clinical relevance, in part due to the inability to fabricate large-scale constructs with physiologically relevant architecture. For this review, a large-scale construct refers to a tissue model with physical dimensions greater than 1 cm³. While smaller constructs provide valuable insight into cellular behaviors and function, scalable macroscale tissues will be essential for functional grafts and physiologically relevant in vitro models. However, shifting from microscale to macroscale bioprinting introduces mechanical and structural challenges, including deformation, poor printability, and mismatched mechanical properties.
Low-viscosity bioinks lack mechanical strength, making large-scale printing difficult with LBB or DBB. This limitation positions EBB as the most suitable modality for printing macroscale cardiac constructs. While high- viscosity bioinks improve shape fidelity upon deposition, soft tissues—including cardiac tissue—often exhibit poor shape retention post-extrusion. The mechanical properties of many bioinks are insufficient to maintain complex geometry, leading to sagging or deformation in unsupported regions.49,99
Additionally, the inherently slower printing speed of EBB—commonly reported in the range of ~5–25 mm/s for cardiac applications—further limits its efficiency in producing high-fidelity, large-scale tissues.49,57,77,88 To address these issues, researchers have developed support bath techniques to improve print fidelity. One such method is the freeform reversible embedding of suspended hydrogels (FRESH), which temporarily suspends soft bioinks in a thermoreversible gelatin bath to stabilize structures during deposition.99 By preventing structural collapse, FRESH enables the fabrication of complex architectures that would otherwise fail under traditional extrusion bioprinting. This approach has enabled the successful bioprinting of small coronary artery models, heart tubes, tri-leaflet valves, ventricle-like constructs, and even a full-sized adult heart model.49,88,99,100
Another strategy for improving structural reinforcement involves multi-material bioprinting. Although tunable hybrid bioinks such as GelMA-PEGDA provide mechanical improvements, printing with separate biomaterials in distinct regions of a construct offers additional advantages. For instance, sacrificial bioinks have been used to provide temporary structural support, allowing for the formation of hollow or intricate geometries that are later cleared. This technique is particularly useful when support bath methods are not feasible.49,101
Until recently, EBB held the sole advantage in multi-material bioprinting due to its ability to integrate multiple print heads, each of which dispenses distinct bioinks to designated regions within a construct. This capability enables the replication of heterogeneous cardiac tissue composition, improving biomimetic accuracy. However, recent advancements in DLP-based LBB have expanded multi-material capabilities, with printers that allow rotating platforms to switch between multiple bioink reservoirs.102 Concurrently, research into photoactive bioinks continues to expand the range of materials compatible with DLP, including ECM, collagen, GelMA, hyaluronic acid (HA), and PEG/PEGDA blends.51,90,103–105
While maintaining structural integrity is critical, excessive stiffness can also negatively impact physiological function. The elastic modulus of the myocardium (~8–17 kPa) not only enables contraction but also plays a role in cardiac differentiation.61 Constructs that deviate significantly from this range may fail to develop properly, integrate with native tissue, or in extreme cases, exacerbate myocardial damage. Thus, tuning stiffness and elasticity through the manipulation of printing parameters—rather than material composition—can enhance structural fidelity while preserving biomechanical properties necessary for functional cardiac constructs. Several bioprinting properties influence construct stiffness. Adjusting pressure, nozzle diameter, and flow rate not only modulates construct stiffness but also impacts cell viability and resolution. Additionally, fine-tuning layer thickness and print infill patterning increase overall stiffness, albeit in a more gradual manner. Izadifar et al.106 demonstrated that changing interstrand distances (300, 900, and 1500 µm) and strand alignment angles (0°, 45°, 90°, and 135°) in printed constructs influences not only elastic modulus but also porosity and electrical conductivity. Optimizing construct porosity is crucial for oxygen and nutrient exchange, highlighting how spatial patterning can be leveraged to enhance both mechanical strength and biological outcomes in engineered tissues.106
2.2.3. Structuring anisotropic cardiac architectures
The electrical and mechanical function of the heart is highly dependent on the anisotropic properties of the myocardium, a complexity that remains a major challenge to replicate. At the cellular level, the alignment of the CMs, which are interconnected by gap junctions, allows rapid, unidirectional electrical signal propagation and synchronized contraction.107 At the tissue level, ECM fibers—predominantly collagen—form structured networks that guide cell alignment, facilitate mechanotransduction, and transmit contraction forces throughout the myocardium. Failure to capture both ECM and myofiber anisotropy can impair electrical conduction and weaken the contraction forces of engineered cardiac tissues.107,108 At the organ level, muscle layers form aligned sheets in a helical arrangement, which gives the ventricles their twisting motion during contraction (Figure 4). Failure to incorporate this architecture can result in lower ejection fractions in chamber models.109

Figure 4. Architecture and anisotropy of cardiac tissue across cellular, tissue, and organ levels. At the cellular level, cardiomyocyte alignment and gap junction connectivity enable unidirectional electrical propagation. ECM fibers and organized myocyte sheets facilitate coordinated mechanical contraction at the tissue level. At the organ level, the helical fiber orientation of multiple layers of the myocardium enables efficient torsional movement during ventricular contraction. Created with BioRender.com. Abbreviation: ECM: Extracellular matrix.
Bioprinting has expanded the ability to control cellular and fiber alignment, bringing engineered tissues closer to replicating the anisotropic nature of native myocardium. Given the unique mechanisms of each modality, researchers have used different approaches to recreate the heart’s architecture based on the individual strengths of different printing modalities.
Recent studies have demonstrated that shear stress-induced alignment combined with nozzle shape optimization and print speed manipulation significantly enhance ECM fiber alignment during EBB. By increasing print speeds to over 2000 mm/min and adjusting the printing pressure, Bera et al.108 directed cells to orient along the printing path, producing patches with aligned ECM fibers that revealed significant improvements in contractility and signal propagation, as evidenced by upregulated expression of contractility genetic markers and intercalated disc formation. Despite successful instances of EBB replicating anisotropy, its ability to fully replicate fine-scale anisotropic features seen in native tissue is severely limited by its low resolution (200–1000 µm).
Both DBB and LBB provide significantly higher resolutions (10–50 µm) compared to EBB. However, DBB’s practical application is restricted by the mechanical properties of its bioink. Due to reliance on low-viscosity hydrogels and cell suspensions, DBB struggles to print large-scale, 3D constructs. This limitation confines its use to thin cardiac sheets and microtissues.110–112
Among LBB techniques, micro-continuous optical printing (μCOP), a subset of DLP achieves even finer resolutions (<10 µm), enabling rapid generation of highly aligned tissue fibers.51,103 Unlike approaches that rely on post-printing cell seeding, μCOP enables direct cellular encapsulation within photopolymerizable hydrogels, ensuring strong initial alignment. μCOP techniques have reported significant enhancement of contractility. Liu et al.103 reported a four- to ten-fold increase in contractile force of aligned constructs compared to 2D-seeded prints.
2.2.4. Integrating vascular networks into bioprinted constructs
To sustain thick cardiac tissues, integrated vascular networks are essential for providing oxygen and nutrients beyond the diffusion limit (200 µm).113 Without perfusable vascularization, engineered constructs are prone to hypoxia and necrosis, restricting the size of viable tissues. Several bioprinting strategies have been developed to incorporate multiple scales of vascular structures, ranging from large perfusable vessels to microcapillary networks (Figure 5).

Figure 5. Vascularization strategies for bioprinting cardiac tissues. (A) Overview schematic of techniques used to incorporate vessels and vascular networks into cardiac constructs, including direct vessel printing, sacrificial bioink-based approaches, and endothelial cell-guided self-assembly. Created with BioRender.com. (B) Hinton et al.99 used direct printing and a 3D MRI scan of a right coronary arterial tree (left) to fabricate large-scale vessels within alginate prints in gelatin support baths (right). (C) Maiullari et al.34 compared cardiomyocyte expression markers TNNI (red) and Cx43 (green) and HUVEC expression vWF (green) in prints using different printing geometries to determine the effect of cellular spatial arrangement on de novo vessel formation. (D) Noor et al.54 created a 3D model of a cardiac patch using sacrificial ink (left) to print a cardiac patch with channels, which form a vascular pattern (right). Panels B, C, and D were adapted from refs. 99, 34, and 54, respectively, under Creative Commons Attribution 4.0 International (CC BY 4.0). Abbreviations: CMs: Cardiomyocytes; EC: Endothelial cell; HUVEC: Human umbilical vein endothelial cell; MRI: Magnetic resonance imaging; OM+CM: Omentum + Cardiomyocytes; vWF: von Willebrand factor.
One approach to integrating vascular networks into bioprinted constructs is to directly print vessels within cardiac constructs. Hinton et al.99 used EBB and FRESH to fabricate coronary arterial trees with internal lumen diameters ranging from 1 to 3 mm and a wall thickness of less than 1 mm. However, when perfusion tests were conducted, they needed to use an external fixture to maintain mechanical stability during the experiment, highlighting the challenges in balancing structural stability with perfusability in directly printed vessels.99 Jia et al.91 used coaxial extrusion to easily create hollow tubes to create vascular of various dimensions. By adjusting the gauging of both the inner and outer needles, they created a diverse range of vessels with inner diameters (400–1000 µm), outer diameters (500–1500 µm), and wall thicknesses (60–280 µm). They proceeded to print an 8 × 9 × 7 mm, 10-layer construct using hollow fibers, producing a completely perfusable construct that demonstrated no rupturing or collapsing.91 The direct printing enables large- scale vessel formation but lacks the resolution needed to form capillary networks, which are critical for cardiac tissues considering every CM has contact with at least one capillary, due to the high demand for nutrients needed for cardiac contraction.16
Another strategy is to print and then remove sacrificial bioink-formed vascular networks leaving behind channels. This approach allows finer-scale vessel formation. Skylar-Scott et al.101 developed the Sacrificial Writing into Functional Tissues (SWIFT) specifically for creating perfusable vascular channels in thick tissues. Using this method, they were able to create channels with a diameter of 400–1 mm within high-density constructs. Lee et al.49 used a similar approach, by embedding and then melting out gelation microparticles in small FRESH-printed collagen discs, creating capillary-sized (~25 μm diameter) pores.
Rather than directly printing vessels or channels, another approach is to promote the natural formation of vasculature through EC patterning and guided self-assembly. Prior to implanting the collagen discs, Lee et al.49 added fibronectin and VEGF to enhance vascularization. When compared to controls, host-derived vessels (8–50 μm) were only observed in printed constructs, demonstrating that bioprinting can guide in vivo vascularization. Maiullari et al.34 used manipulation of cell spatial arrangements to enhance vascularization. Using an alternating layer arrangement of iPSC-CM and HUVECs, they were able to generate vessel-like structures (~150 μm lumen diameter) more effectively than with other spatial arrangements.
Table 3 presents a selection of representative cardiac bioprinting studies that employ different modalities, materials, and construct types. While not exhaustive, these examples illustrate key advances and diversity in cardiac bioprinting approaches.
Table 3. Modalities, materials, and construct types used in bioprinting of cardiac constructs
| Modality | Design | Biomaterials | Cells | Key outcomes | References |
|---|---|---|---|---|---|
| Extrusion | Engineered cardiac tissue | GelMA Laminink-521 | iPSC-CMs | Developed a patient-derived bioprinted patch for CHD modeling | 25 |
| Extrusion (coaxial) | Engineered cardiac tissue | Alginate, PEG- fibrinogen, CaCl2 solution | iPSC-CMs, HUVECs | Coaxial bioprinting of vascularized cardiac patches with endothelialized microchannels | 34 |
| Extrusion | Cardiac patch | Omentum-derived dECM hydrogel | iPSC-CMs, iPSC-ECs | Created for the first fully personalized, thick, and perfusable patches | 54 |
| Extrusion | Engineered cardiac tissue | Omentum-derived dECM hydrogel | iPSC-CMs, iPSC-ECs | Developed thick vascularized cardiac tissues with perfusable triaxial lumens | 54 |
| Light (DLP) | Engineered cardiac tissue | GelMA | hPSC-pacemaker cells | Bioprinted a pacemaker-like construct capable of pacing a hPSC-CM laden “Mini Heart” model | 89 |
| Light (DLP/μCOP) | Engineered cardiac tissue | GelMA, PEGDA, H54AGM | iPSC-CMs, human CFs | Alignment cues used to develop cardiac microtissues for high-throughput drug testing | 51 |
| Extrusion | Engineered heart tissue | Collagen, Matrigel® | iPSC-CMs, CFs | Printed a heart tube model | 88 |
| Extrusion | Engineered heart tissue | Fibrinogen | hiPSC-CMs, hiPSC-ECs, primary adult ventricular human CFs | Aligned, high-cellular-density cardiac tissue produced in highly reproducible format (24-well plate) | 53 |
| Extrusion | Engineered cardiac tissue | GelMA, collagen | iPSC-CMs from healthy and CPVT patients | First functional printed CPVT patient-specific model | 32 |
| Light (DLP/ μCOP) | Engineered cardiac tissue | GelMA | hESC-CMs | GCaMP3-hESC-CMs* printed into patterned hydrogels for simultaneous force and calcium readouts | 90 |
| Extrusion | Engineered heart tissue | GelMA/ColMA | hiPSCs → hiPSC-CM | First macroscale, perfusable two-chambered heart model | 23 |
| Extrusion | Engineered Heart tissues | Collagen/hyaluronic acid | hiPSC-CMs | Directly bioprinted beating ring and ventricle constructs | 24 |
| Extrusion | Engineered cardiac tissue | Collagen | iPSC-CMs | Bioprinted high-fidelity heart components across multiple scales: perfusable vascular networks, tri-leaflet valves, left ventricle model, neonatal-scale whole human heart (two-chambered) | 49 |
| Extrusion | Valve | PEGDA, alginate | Porcine aortic valve interstitial cells | Printed anatomically accurate, and mechanically heterogeneous aortic valves | 77 |
| Extrusion (coaxial) | Vascular constructs | GelMA, PEGTA | HUVECs, iPSC-derived ECs | Single-step direct bioprinting of perfusable, multi-layered vascular constructs with tunable lumen geometry | 91 |
| Extrusion | Engineered cardiac tissue | Fibrin, gelatin | iPSC-CMs, CFs | Demonstrated connexin 43 mediated electrical coupling and myocardium- matched stiffness via dual crosslinking | 67 |
| Extrusion | Engineered cardiac tissue | GelMA–alginate | hiPSC-CMs, HUVECs | Developed an anisotropic, endothelialized myocardium-on-a-chip model for drug cardiotoxicity evaluation | 35 |
*GCaMP3 is a calcium indicator.
Abbreviations: CaCl2: Calcium chloride; CFs: Cardiac fibroblasts; CHD: Congenital heart disease; ColMA: Collagen methacrylate; CPVT: Catecholaminergic polymorphic ventricular tachycardia; dECM: Decellularized extracellular matrix; DLP: Digital light processing; GelMA: Gelatin methacrylate; H54AGM: Glycidyl methacrylate-hyaluronic acid; hESC-CMs: Human embryonic stem cell-derived cardiomyocytes; hiPSCs: Human induced pluripotent stem cells; hPSC: Human pluripotent stem cells; HUVECs: Human umbilical vein endothelial cells; iPSC-CMs: Induced pluripotent stem cell-derived cardiomyocytes; iPSC-ECs: Induced pluripotent stem cell-derived endothelial cells; μCOP: Micro-continuous optical printing; PEG: Poly(ethylene glycol); PEGDA: Poly(ethylene glycol)-diacrylate; PEGTA: 4-arm poly(-ethylene glycol)-tetra-acrylate.
2.3. Post-processing
Post-processing is essential for transforming initial prints into structurally stable and functionally mature tissue. Without post-printing modifications, engineered tissues often lack mechanical strength, mature electromechanical activity, and sufficient vascularization for physiological function. Common post-processing strategies include crosslinking for mechanical stabilization and functional conditioning to promote cellular maturation.
Crosslinking, the process of forming bonds within a bioink polymer network, is a critical step in bioprinting to maintain structural integrity and enhance mechanical stability. Choosing a crosslinking strategy influences the mechanical properties, biocompatibility, and degradation of the printed construct. Crosslinking can be divided into physical and chemical techniques, with the most commonly used techniques including photocrosslinking, ionic crosslinking, thermal, and enzymatic crosslinking.114–117
Physical crosslinking techniques—such as ionic and thermal crosslinking—use changes in external stimuli to induce non-covalent bonding in the polymer chains of the biomaterial. Ionic crosslinking relies on multivalent cations to induce gelation. The most common ionic crosslinker ion in bioprinting is calcium, which is frequently applied in alginate-based bioinks. 63,115 Other ionic crosslinkers, including barium chloride (BaCl2) and zinc chloride (ZnCl2), have been tested for their influence on the mechanical and biological properties post-crosslinking; however, BaCl2 had biocompatibility concerns while ZnCl2 resulted in cytotoxic effects.118 Other calcium-based crosslinkers include calcium sulfate (CaSO4) and calcium carbonate (CaCO3), which offer slower Ca²+ release potentially leading to more uniform and controlled crosslinking but are often not utilized because CaCl2 offers a faster, simpler, and well-established solution.116,119 Although rapid and highly biocompatible, CaCl2-crosslinked constructs tend to be mechanically weaker, limiting their long-term structural stability.
Thermal crosslinking involves temperature-dependent gelation of thermosensitive polymers such as collagen, gelatin, GelMA, and Pluronic F127.117,120 While operationally straightforward, effective thermal crosslinking can be challenging due to a lack of ability to precisely control the degree of crosslinking through temperature shifting alone. Furthermore, some materials can take minutes or hours to thermally crosslink but seconds with alternative methods. The temperature at which a biomaterial crosslink is based not only on the material type but also on other factors such as concentration. For example, the percentage at which GelMA is reconstituted determines the temperature needed to induce gelation. 10% GelMA—commonly used in bioink formulations—gels around 23°C, while GelMA of higher concentrations, such as 15%, gels at 27°C.120 Thermally crosslinked constructs exhibit low mechanical strength, which explains their relatively short-term stability. However, this short-term stability is often taken advantage of to produce sacrificial bioinks and temporary support baths/structures as previously described.53,54,88,100,101 In general, physical crosslinking techniques are weaker than chemical techniques and are often reversible.
Chemical crosslinking, on the other hand, forms covalent bonds between polymer chains, which are stronger and often permanent. These techniques are particularly important for long-term structural stabilization. Photocrosslinking, the most common crosslinking strategy employed in cardiac bioprinting, uses light exposure to initiate polymerization in bioinks containing photoinitiators, making it a widely applicable technique across EBB, LBB, and DBB. The most common photoinitators in bioprinting are Irgacure-2959, lithium phenyl-2,4,6-trimethylbenzoylphosphi-nate (LAP), and eosin Y. Irgacure and LAP are UV l-activated crosslinkers, typically excited at 365 or 405 nm, enabling rapid gelation and tunable mechanical properties.114 However, UV light can be cytotoxic, reducing cell viability within printed constructs. As an alternative, visible light crosslinking systems—such as Eosin Y in combination with initiator triethanolamine, and catalyst 1-vinyl-2 pyrrolidinone—have been developed to minimize cell damage while maintaining effective crosslinking.67,121
Enzymatic crosslinking is another common type of chemical crosslinking that uses enzymes—commonly, transglutaminase and horseradish peroxidase—to induce gelation in a gentler but highly specific reaction. Although it has higher biocompatibility, is tunable and allows in situ crosslinking, its slow gelation and sensitivity to environmental conditions such as oxygen, pH, and temperature can be limiting.115,122,123
Some researchers are using hybrid crosslinking approaches to improve structural fidelity without sacrificing biocompatibility.67,91,122,123 Basara et al.122 used photocrosslinking and enzymatic crosslinking to model myocardial infarct boundaries where the stiffness of healthy cardiac tissue (8–12 kPa) and scar tissue (>150 kPa) meet. UV light was used to rapidly crosslink a GelMA–MeHA–dhECM bioink, which under normal conditions tends to be mechanically weak after printing. After the initial crosslinking, they use microbial transglutaminase (mTGase) to further crosslink the gels, resulting in tissues with stiffness ten times greater than GelMA controls, allowing them to mimic both healthy and fibrotic tissue.122 Contrary to Basara et al.’s dual techniques that used both crosslinking techniques after printing, it is possible to use crosslinking prior to printing. Budharaju et al.123 combined ionic and enzymatic crosslinking to enhance shape fidelity, printability, and mechanical strength of alginate–fibrinogen-based myocardial patches. First, CaCl2 was used to pre-crosslink the alginate prior to printing, resulting in gelation suitable for extrusion without inducing nozzle clogging and significant improvement of shape fidelity and print resolution. Once the constructs were printed, additional CaCl2 was used to fully crosslink the alginate along with thrombin which was used to crosslink the fibrinogen. This dual crosslinking completed the mechanical stabilization of the constructs, with hybrid crosslinking demonstrating slower degradation rates than single crosslinked controls.123
Although crosslinking strategies ensure long-term mechanical integrity, structural stabilization alone is insufficient for generating fully functional cardiac tissues. Physiological conditioning is essential for CM maturation, synchronized electrical activity, and functional vasculature. To achieve this, researchers employ mechanical loading, electrical stimulation, and perfusion techniques to replicate the native cardiac environment and enhance long-term tissue viability.
The heart is constantly subjected to mechanical stress due to continuous myocardial contraction, which plays a critical role in CM development and maturation.124In vitro, tissue stretching or compression has been used to simulate these mechanical forces, leading to increased sarcomere lengths, larger cell area, elongation, improved calcium handling, and greater expression of sarcomere, ion channel, and maturation-related genes.125,126 Furthermore, mechanical stimulation enhances CMs alignment, showing cyclic stress is more effective than static stress, likely due to its rhythmic nature, matching that of the heart.127
Electrical stimulation is also a widely used strategy for enhancing the maturation of engineered cardiac tissues. Electrical stimulation has been shown to accelerate sarcomere organization, enhance calcium handling, and upregulate electrophysical gene expression.128,129 Notably, a gradual increase in stimulation is more effective over time than maintaining a fixed pacing rate.129,130 Emerging approaches, such as mechanical and electrical stimulation, advanced bioreactor systems, and machine learning-based bioink optimization, show promise for improving cell maturation, vascularization, and tissue endurance.125,131
Beyond mechanical and electrical conditioning, pulsatile perfusion plays a dual purpose in post-processing. It enhances cell survival by continuously supplying nutrients and removing metabolic waste while simultaneously delivering biomechanical cues to stimulate CM maturation. Additionally, flow-induced shear stress promotes EC alignment, further enhancing vascularization.131,132
While each of these conditioning strategies has been shown to enhance tissue function individually, their combined application can have a compounding effect, significantly improving overall tissue maturation. As a result, bioreactors are becoming indispensable in post-processing, allowing for simultaneous integration of multiple conditioning techniques, effectively replicating the complex and dynamic physiological environment of the native heart.131–133
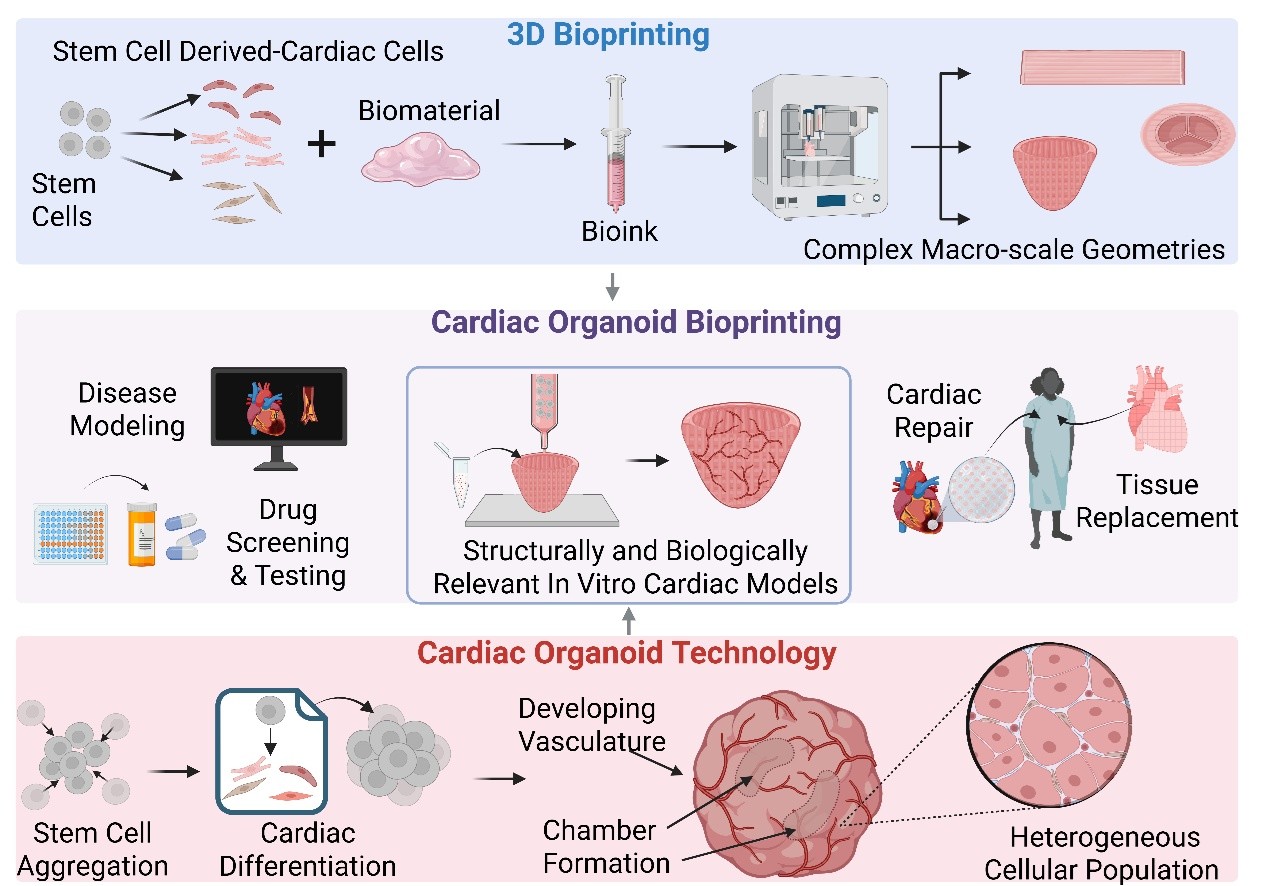
Bioprinting has enabled the fabrication of diverse structured cardiac tissues, advancing in vitro modeling beyond 2D culture, animal models, and conventional 3D cultures. However, while bioprinting allows precise spatial control over tissue structures, it does not fully capture the developmental self-organization inherent to natural heart development. Cardioids, or cardiac organoids, represent a promising model that leverages biological self-organization to recapitulate aspects of early heart development. These models recapitulate key aspects of early heart formation, including cellular diversity and functional mimicry, providing valuable insights into cardiac biology.12–14 Integrating bioprinting with cardioid technology may provide a hybrid approach, combining biological self-organization with engineered structural guidance to enhance scalability, vascularization, and maturation. The following section will explore the fundamental characteristics of cardioids, their applications in cardiac research, and their current limitations.
3. Cardioids: features, applications, and limitations
Cardioids, or cardiac organoids, are self-organizing, 3D heart tissues that model aspects of early heart development in vitro. Historically, the term “cardiac organoid” was first used in a 2001 study by Zimmerman et al.,11 who cast neonatal myocytes and collagen type I into circular molds to generate a contractile cardiac construct.
The observation of self-organization post-fabrication and the presence of non-myocyte populations led the authors to label their construct as “organoid-like.”11 Broadly, the term “organoid” has been used loosely and interchangeably with several other terms referring to 3D cultures—such as microtissues, assembloids, and spheroids, and engineered heart tissues—yet there are universally agreed upon features that all organoids, including cardioids possess.7,134 In this review, we define cardioids as possessing four key characteristics: self-organization, complex 3D architecture, cellular diversity, and functional mimicry (Figure 6).

Figure 6. Schematic depicting four key features of cardiac organoids. Cardiac organoids exhibit: (1) self-organization, where stem cells autonomously aggregate and differentiate to form tissue structures; (2) cellular heterogeneity, with multiple cardiac cell types contributing to cardioid function; (3) complex 3D architecture, including heterogeneous layer formation, primitive vascularization, and chamber-like cavity formation; and (4) functional mimicry, as demonstrated by synchronous beating and responsiveness to external stimuli such as drug exposure. Created with BioRender.com.
3.1. Cardioid features
A defining hallmark of cardioids is their intrinsic ability to self-organize into 3D structures without the need for scaffolding or external patterning. Typically, cardioids are formed via directed differentiation of PSCs, allowing cardioids to develop diverse cellular populations through a process that closely mimics early embryonic development.12 This process enables cardioids to develop a 3D architecture that closely resembles the in vivo organization of embryonic cardiac tissue. This includes multiple organized layers of a diverse population of cells, including CMs, ECs, and CFs, as well as surrounding ECM components that help guide self-organization.135–143
In contrast, spheroids mainly refer to simple, scaffold- free aggregates of a single cell type, while assembloids describe the fusion of organoids with other tissues or additional organoids.7,144 Engineered heart tissues are another type of in vitro model that are sometimes referred to as cardioids. Although engineered heart tissues can exhibit multi-dimensional architectures, they are typically fabricated using forced assembly techniques involving pre-patterning, forced aggregation, molds, scaffolds, or biomaterial supports.7 Additionally, most engineered heart tissues utilize pre-differentiated cardiac cells, bypassing the self-organization of PSCs. However, because these cells are derived independently, the constructs lack the intrinsic signaling interactions between myocytes and non-myocytes that coordinate developmental cell fate decisions, leading to incomplete cellular differentiation and limited maturation. This approach necessitates separate differentiation protocols for each major cell type (e.g., Wnt/β signaling for CMs, VEGF/BMP for ECs, and FGF/TGF-βi for CFs).21,22,38,39,46,47 These models tend to exhibit more adult-like properties and, as a result, are often used for electromechanical models of the heart. Additionally, this method often fails to incorporate minor cardiac cell types due to the lack of well-defined differentiation protocols.20 Furthermore, many engineered heart tissues have simple designs—such as strips, rings, patches, tubes, and single chambers—that often recapitulate only one cardiac property rather than replicating the full architectural complexity and functionality of the developing heart.26–31,43–45,50,52,129
Unlike traditional in vitro cardiac models, cardioids recapitulate key morphogenetic events observed in early cardiac development including chamber formation, outflow tract emergence, and lineage specification.135–138 These processes are absent in traditional 2D monolayer cultures and only partially represented in other 3D models. Many cardioids exhibit electromechanical functions including spontaneous contractions and electrical conduction, demonstrating primitive but physiologically relevant early cardiac activity. Additionally, some cardioids display rudimentary aspects of valvular and conduction system development, reminiscent of embryonic stages of the heart. These features make cardioids valuable models for studying cardiac morphogenesis, congenital heart defects, and disease pathophysiology—topics that will be further explored in future sections.7,14,134
While traditional cardioids form through self-assembly, engineering strategies can be used to refine and support self-organization, enhancing reproducibility, and structural complexity while still preserving the intrinsic mechanisms that drive heart formation. These hybrid approaches blur the line between cardioids and engineered heart tissues, but if self-organization remains a defining feature post-printing they can still be classified as cardioids.
3.2. Cardioid formation
Early cardiac differentiation protocols relied on embryoid bodies-based differentiation, ESCs or iPSCs spontaneously aggregated in suspension cultures, forming 3D clusters.145 These aggregates developed multiple germ layers including the cardiac mesoderm without external signals, and within several days a small subset of cells would begin spontaneous contraction. While this approach was cost-effective, scalable, and required minimal intervention, it yielded highly heterogeneous populations with low CM efficiency, limiting its applicability for cardiac-specific models.145,146
To improve cardiac cell, yield, and uniformity, differentiation strategies shifted toward directed monolayer-based approaches, which became increasingly efficient at generating CMs.21,22 However, this came at the cost of replicating 3D cellular interactions and incorporating essential non-myocyte populations.20 Since heterogeneous cellular composition is critical for functional cardiac tissues, interest in 3D differentiation models has re-emerged, but with greater emphasis on directed differentiation strategies.16
Self-assembly-based cardioid formation typically begins with PSCs being cultured in suspension culture, where intrinsic signaling and cell–cell interactions promote cellular aggregation. Suspension culture environments—most commonly ultra-low attachment plates—provide a scaffold-free setting in which cells can self-organize.136–138,142,143 Although scaffolding is not as common, in cardioid development, some researchers chose to encapsulate their cardioid in a supporting ECM such as collagen or Matrigel®, to provide additional biological cues for differentiation, better mimicking the 3D aspect of the native cardiac microenvironment.135,142
Comparable to traditional monolayer approaches, the most common method for cardioid differentiation involves modulating the Wnt/β-catenin pathway with small molecules. Activation of the pathway—typically with Chir—triggers mesodermal induction, where PSCs first differentiate into mesodermal progenitors and subsequently into cardiomesodermal progenitors.83 Additionally, growth factors such as bone morphogenetic protein 4 (BMP4), activin A, Wnts, and FGFs are utilized to help generate cardiac mesodermal cells. From the cardiomesoderm, various progenitor cells spawn the major cardiac cells: CMs, cardiac ECs, and CFs.7,20 VEGF—a key player in vascular specification, FGF and TGF-βi—commonly used for the generation of epicardial cells, are common growth factors used in cardioid protocols.7,12
Cardioid research gained traction in 2021, with several overlapping studies demonstrating the emergence of cardiac structures through precise modulation of Wnt, BMP, and FGF signaling (Table 4). Hofbauer et al.138 achieved chamber-like structures with endocardial and epicardial layers and clarified the relationship between HAND1, a transcription factor linked to developmental heart chamber defects, and chamber formation. Lewis-Israeli et al.136 used three-step Wnt modulation (activation/inhibition/activation) to guide atrioventricular specification, generating cardioids with distinct atrial and ventricular CMs (Figure 7A). In 2023, Volmert et al.143 used fatty acid supplementation to induce a metabolic shift from glycolysis to fatty acid oxidation (Figure 7B). They observed the formation of distinct atrial and ventricular regions, endogenous anterior–posterior patterning following retinoic acid exposure, and the emergence of valvular and conductance cells for the first time within a cardioid.
Table 4. Highlights from cardioid publications in recent years
| Culture method | Cells/tissue | Biomolecules & growth factors | Features | References |
|---|---|---|---|---|
| Ultralow-attachment 96-well plate, aggregates embedded in Matrigel® droplets | hESCs and hiPSCs → CMs, endocardial-like cells, foregut endoderm, septum transversum-like mesenchyme | Chir (7.5 µM, day 0), IWP2 (5 µM, day 3–5) | Chamber-like structures, formation of primitive heart fields, endocardial lining developed, vascular-like endothelial organization, primitive heart field emergence, foregut endoderm regions | 135 |
| Ultralow-attachment 96-well plate | hESCs & hiPSCs → CMs, endocardial-like cells, epicardial-like cells | Chir, BMP4, activin A, FGF2, retinoic acid, LY294002, IWR-1 | Chamber-like structures, epicardial and endocardial-like layers, fibrotic response upon cryoinjury | 138 |
| Ultralow-attachment 96-well plate | hESCs & hiPSCs → atrial & ventricular CMs, epicardial cells, endocardial cells, ECs | Chir, C-59, BMP4, activin A | Distinct atrial and ventricular regions, myocardial cavities, pre-gestational diabetes model | 136 |
| Ultralow-attachment 96-well plate | mESCs → CMs, early heart field progenitors, vascular-like ECs, endocardial-like cells | Chir, bFGF, VEGF, ascorbic acid | Formed gastruloids, early heart field-like structures, multi-lineage interactions; first heart field patterning, early vascular-like structures, cardiac crescent, and heart tube-like structure gut-like tube formation | 137 |
| Tissue culture plate (iPSCs → mesendoderm progenitors), 6-well low-attachment tissue culture plates ( mesendodermal spheroids) | hiPSCs → CMs, mesodermal derivatives, gut-like endodermal cells, epicardial-like cells, smooth muscle cells | Chir, IWP2 FGF, retinoic acid, ascorbic acid | Atrial/nodal CMs development, cardiac (mesoderm), and gut (endoderm) tissue formation, epicardial layer and contractile smooth muscle cells (peristalsis behavior) development | 139 |
| AggreWell™ 800 | hESCs & hiPSCs → CMs, epicardial-like cells, foregut- derived hepatic progenitors, and mesenchymal cells | Chir, retinoic acid, BMP4, IWP-4 | Co-culture of CM aggregates and PE/STM foregut organoid formed epicardium-myocardium organoids, epicardial-like layer surrounding myocardium layer, ECM deposition at the periphery | 140 |
| Six-well plates on orbital shaker | hiPSCs → CMs, epicardial cells CFs, vascular endothelial cells, neural & mesodermal derivatives | Chir, FGF2, VEGF, ascorbic acid, IGF, HGF | Thin-walled, dilated chamber-like structures, elaborate neuron structural network, outflow tract-like structure | 141 |
| Ultralow-attachment 96-well plate, collagen embedment post-differentiation | hiPSCs → ventricular CMs, pacemaker cardiomyocytes, epicardial cells, epicardium- derived cells | BMP4, bFGF, retinoic acid, IWP2, VEGF | Generated epicardioids, vessel-like structures, ventricular myocardium and epicardium layers | 142 |
| 96-well ultra-low-attachment plates | hESCs & hiPSCs → ventricular and atrial CMs, valve cells, proepicardial-derived cells, epicardial cells, stromal cells, cardiac progenitor cells, conductance cells, ECs | Chir, BMP4, activin A, maturation media (oleic acid, linoleic acid, palmitic acid, L-carnitine), Wnt-C59 | Large atrial & ventricular chambers, retinoic acid gradient-driven anterior–posterior patterning, post-natal metabolic activity | 143 |
Notes: LY294002 is a protein inhibitor; IWP2, IWP-4, IWR-1, and Wnt-C59 are Wnt signaling pathway inhibitors.
Abbreviations: bFGF or FGF2: Basic fibroblast growth factor; BMP4: Bone morphogenetic protein 4; CFs: Cardiac fibroblasts; Chir: CHIR99021- glycogen synthase kinase 3 beta inhibitor; CMs: Cardiomyocytes; ECs: Endothelial cells; ECM: Extracellular matrix; FGF: Fibroblast growth factor; hESCs: Human embryonic stem cells; HGF: Hepatocyte growth factor; hiPSCs: Human induced pluripotent stem cells; hPSC: Human pluripotent stem cells; HUVECs: Human umbilical vein endothelial cells; IGF: Insulin-like growth factor; iPSC-CMs: Induced pluripotent stem cell-derived cardiomyocytes; iPSC-ECs: Induced pluripotent stem cell-derived endothelial cells; PE/STM: Pro-epicardium/septum transversum; TGF-β: Transforming growth factor beta; VEGF: Vascular endothelial growth factor.

Figure 7. Examples of different approaches to cardioid formation. (A) Lewis-Israeli et al.136 utilized a three-step WNT modulation protocol (left) to generate cardioids with distinct atrial and ventricular regions. A microscopic image of a day 15 cardioid (right) displays expression of DAPI (blue), mature ventricular MYL2 (green), and atrial MYL7 (red). Scale bar = 500 μm. (B) Volmert et al.’s schematic143 (left) illustrates their differentiation protocol and fatty acid supplementation strategy. Immunofluorescence imaging of a day 30 cardioid (right) cultured in the advanced maturation media (EMM2/1) demonstrates distinct atrial and ventricular regions, as evidenced by expression of atrial marker NR2F2 (green), ventricular marker MYL3 (red), and DAPI (blue). Scale bar = 200 μm. (C) Branco et al.’s schematic140 (left) shows a co-culture strategy with foregut-derived organoids, and the microscopic image (right) displays a day 15 epicardium-myocardium cardioid stained for mature ventricle marker MLC2V (green), atrial/immature ventricle marker MLC2A (red), and DAPI (blue). Scale bar = 100 μm. Panels A, B, and C were adapted from refs. 136, 143, and 140, respectively, under Creative Commons Attribution 4.0 International (CC BY 4.0).
Proper cardiac morphogenesis depends on mechanical cues and paracrine signaling from neighboring tissues, such as the endothelium and foregut. Several studies have attempted to incorporate the generation of multiple germ layers to better mimic early heart development. Rossi et al.137 developed gastruloids, which mimic gastrulation—the embryonic stage where the three germ layers form. These pre-cardiac models expressed both first and second heart field progenitors and contained a primitive gut-like tube, vascular network, and endocardial-like layer. Building on this concept, Drakhlis et al.135 generated heart-forming organoids that included early heart and foregut tissues, comprising mesodermal layers lined with endocardial-like cells and splanchnic mesoderm, along with gut endodermal regions. Silva et al.139 further demonstrated that gut–heart interactions enhanced mesodermal signaling, promoted advanced structural and functional maturation of CMs, induced atrial/nodal subtype specification, and observed the development of contractile smooth muscle-like cells that exhibited peristalsis-like movements. Collectively, these studies illustrate that multi-germ layer interactions enhance CM maturation and promote the development of specialized cardiac subtypes, thereby improving the physiological relevance of cardiac organoids for studying complex processes beyond early development.
Beyond modeling early cardiac development, some studies have explored multi-organ interactions to better understand cardiogenesis in a broader embryonic developmental context. Olmsted and Paluh141 created elongating multi-lineage organized cardiac gastruloids, which exhibited chamber-like structures, outflow tractlike patterning, as well as an elaborate neural network, offering insight into cardio-neuro interactions during cardiogenesis and neurogenesis.
Complex cardioid systems have been achieved through co-culture approaches. Branco et al.140 developed pro- epicardium/septum transversum foregut organoids which contained both epicardial progenitors and hepatic foregut derivatives, reflecting the developmental link between the heart and liver. When combined with CM aggregates, epicardium-myocardium organoids formed, in which an epicardial-like layer surrounded the myocardium, influencing cardiac maturation and leading to ECM deposition (Figure 7C).140
3.3. Applications
Cardioid’s ability to mimic key morphogenetic processes, including chamber formation, metabolic transitions, and electrophysiological activity, make them well-suited for investigating both normal and pathological cardiac function. Furthermore, cardioids’ 3D architecture, cell diversity, and functional properties enable a more comprehensive assessment of drug-induced effects, ranging from electrophysiological disturbances to tissuespecific toxicity and regenerative potential.
Cardioids have provided key insights into developmental defects, injury responses, and cardiac regeneration. Hofbauer et al.138 observed physiologically relevant responses, including severe necrosis, continued contractions, fibroblast migration toward the injury site, and ECM accumulation, after subjecting their cardioids to cryoinjuries, mirroring aspects of both normal wound healing and pathological fibrosis. Voges et al.147 also used cryoinjury to assess the regenerative capacity of early- stage human heart tissue, demonstrating high rates of CM proliferation and recovery of cardioid contractility, suggesting an innate regenerative potential in immature cardiac tissue. Lewis-Israeli et al.136 investigated the effects of pregestational diabetes on cardiac development by exposing heart-forming organoids to hyperglycemic conditions, resulting in hypertrophic phenotypes, reduced mitochondrial content, dysfunctional lipid metabolism, and structural disorganization.
Cardioids have also proven particularly effective in identifying off-target effects of non-cardiac drugs. Ondansetron (Zofran®), an anti-nausea medication commonly prescribed during pregnancy, has been linked to congenital heart defects.148 When cardioids were exposed to Zofran® at increasing concentrations (1, 10, and, 100 μM), ventricular CM populations were significantly reduced in a dose-dependent manner, while atrial CMs remained largely unaffected. At the highest concentration, cardioids exhibited structural disorganization and loss of chamber wall definition.143 Similarly, cardioids have been used to investigate the cardiotoxic effects of doxorubicin, a widely used anticancer drug. Post-myocardial infarction organoids exposed to doxorubicin (0.1–50.0 µM) exhibited decreased contractile function, increased cell death, and severe structural disruptions in CMs, in a dose-dependent manner, with infarcted cardioids showing side effects at lower dosages than non-infarcted control organoids, consistent with the clinical observations of doxorubicin- induced cardiomyopathy in patients with pre-existing cardiovascular conditions.149,150 In the same study, cardioids were used to investigate JQ1, a potential heart-failure drug candidate. Infarcted organoids treated with JQ1 exhibited reduced fibrosis and improved synchronized beating, suggesting its potential as a therapeutic for post-MI recovery.149
These studies highlight the value of cardioids as physiologically relevant models for disease modeling, regenerative medicine, drug safety testing, and therapeutic screening. Their small size—typically ranging from 1 to 2 mm—enables high-throughput screening, optical accessibility, and efficient nutrient diffusion, making them well-suited for developmental and pharmacological studies. However, this microscale also imposes limits on vascularization, maturation, and scalability, which restrict their use in modeling adult physiology/pathology or serving as implantable grafts. As cardioid platforms continue to evolve, integrating them with bioengineering technologies such as bioprinting may help overcome these limitations and expand their applicability to more translational contexts.
3.4. Limitations
While cardioids have advanced our ability to model human cardiac development and disease, several challenges hinder their clinical and translational potential. These include limited tissue maturity, diffusion constraints due to the absence of vasculature, batch variability, and scalability issues.8 Overcoming these challenges will be essential for advancing cardioids in disease modeling, drug discovery, and regenerative medicine.
As with all engineered cardiac tissues, achieving maturity remains one of the most pressing challenges.8 PSC-derived CMs often resemble fetal cells rather than adult CMs, exhibiting small, round morphology, unorganized sarcomeres, few mitochondria, glycolysis-dependent metabolism, asynchronous contractility, and immature calcium-handling properties.124 In contrast, mature CMs exhibit elongated morphology, aligned and organized sarcomeres, abundant mitochondria, oxidative metabolism, synchronized contractions, and efficient calcium handling—features essential for adult-like cardiac tissue function. This immaturity within cardioids not only restricts their clinical applicability but also limits their ability to model adult-onset diseases or assess drug responses in fully developed tissues. Previous studies with bioprinted constructs have demonstrated that strategies such as bioreactors, physical conditioning, and co-culture techniques are necessary to enhance tissue maturation. Thus, these strategies are expected to be critical for improving cardioid maturation. Physical conditioning of increasing intensity was demonstrated to be effective at encouraging an adult-like maturation state of early iPSC-derived CMs if initiated soon after spontaneous contractions began.129 Given the developmental nature of cardioids, implementing these strategies may be particularly impactful for enhancing maturity. The development of a fully perfusable vasculature is crucial for advancing current cardioid toward more physiologically relevant models. Current cardioid models exhibit disorganized capillary structures and lack defined arteries, arterioles, veins, and venules. As a result, they rely on passive diffusion for oxygen and nutrient transport, restricting their size to a microscale range.135,136,143,151 Additionally, the lack of functional vasculature increases the risk of necrotic core formation in larger constructs. Thus, vascularization strategies must be optimized so that cardioids can be successfully scaled up. This presents a unique challenge, as the heart has the highest oxygen demand of any organ, requiring a dense vascular network that is difficult to replicate in vitro.33
The self-organizing nature of cardioids results in batch-to-batch variability, which poses a challenge for consistent structural and function outcomes. Small variations in culture conditions can lead to significant differences between and within batches, posing challenges for standardization, regulatory approval, and clinical translation. Additionally, current methods for generating cardioids are not high-throughput, limiting their use in large-scale pharmaceutical screening. Emerging fabrication strategies, including bioprinting, offer promising solutions to address these challenges by improving scalability, reproducibility, and vascularization.
4. Progress toward cardioid bioprinting
Organoid bioprinting is an emerging technology that has successfully yielded functional bioprinted organoids for many organ systems including the brain, kidney, intestine, liver, lungs, and pancreas151–156. Yet, progress in cardioid bioprinting lags behind other organs due to the mechanical complexity required to support continual contraction, unidirectional force transmission, and synchronized electrical activity, as well as the heart’s high metabolic demand for oxygen and nutrients. 157,158 In this review, we consider two approaches to cardioid bioprinting: (i) using bioprinting strategies to print individual organoids and (ii) using cardioids as the building blocks to print cardiac constructs. In this section, we will discuss the progress in cardioid bioprinting toward addressing the challenges of cardiac tissue engineering discussed in section 2.2, as well as how bioprinting addresses the limitations of standard cardioids.
One major challenge in cardiac tissue engineering is achieving heterogeneous physiological cellular density. Cardioids naturally achieve these densities across multiple cardiac cell types; however, their average diameter is typically only 1–2 mm.135,136,143,151 Bioprinting, as we have seen, can easily produce macroscale cardiac constructs capable of maintaining structural fidelity. Furthermore, there has been recent progress in supporting the printing of PSCs, which are the predominant cells used to generate organoids, though their application is still rather limited.159 Kupfer et al.23 used this approach to develop human chambered muscle pumps (HCHAMPs)—3D EBB-printed chambered cardiac constructs. To support both iPSC encapsulation and CM differentiation, they optimized a GelMA/collagen methacrylate bioink supplemented with native cardiac ECM proteins. The HCHAMPs featured a macroscale (1 cm long), two-chambered structure with inlet and outlet vessels—enabling perfusion, pressure/ volume dynamics, continuous action potentials, and responsiveness to pacing and drug stimuli. However, the HCHAMPs lacked cellular heterogeneity, only containing CMs, which limited their ability to replicate CMs and non-myocyte interactions and develop vasculature.23 Expanding on the novelty of this approach by utilizing cardioid differentiation protocols, instead of CM-only protocols, could simultaneously address many cardiac engineering challenges, including the lack of macro-scale cardioids, the lack of cellular diversity of printed constructs, and limited functional complexity of both cardioids and printed cardiac constructs. However, larger constructs with higher densities exacerbate the need for vascularization.
Another challenge is integrating vascular and anisotropic organization without disrupting the self-organization behavior critical to cardioid development. Direct printing, indirect formation through sacrificial techniques, and encouragement of de novo formation of vasculature have all been used to create vessels and vascular networks in printed constructs, but standard cardioids rely solely on self-organization of vessel-forming cells such as ECs and VSMCs. Furthermore, cardioids rely on self-organization to form the anisotropic organization of cardiac cells and ECM necessary to carry out cardiac-specific tasks such as electrical signal propagation, contraction, and efficient ejection throughout chambers, while this organization must be manually achieved in printed constructs. Hybrid approaches have been developed to combine the manual organization of cells and ECM without overriding self-organization but instead enhancing it. Bioprinting-assisted tissue emergence (BATE) developed by Brassard et al.151 is a novel organoid bioprinting technique that involves the usage of a syringe-based extrusion printer coupled to a microscope and a manually controlled stage to manipulate cell spatial position within ECM, allowing for the printing of centimeter-scale human intestinal, connective, and vascular tissues with complex branched vasculature and luminal structures. If combined with directed differentiation, BATE and similar approaches could provide finer spatial control over cardiac tissue organization, potentially enabling the recreation of cardiac structures such as valves, coronary arteries, and defined atrial and ventricular chambers while allowing the self-organization of the appropriate cells toward the pre-formed structures. Furthermore, combining BATE with the second bioprinting approach of using pre-formed cardioids could further enhance self-organization.
Ahrens et al.160 embedded anisotropic organ building blocks (aOBBs) within a gelatin-fibrinogen bioink to print cardiac macrofilaments and tissue sheets. Within the cardiac sheets, the aOBBs aligned according to the printing patterns, which included linear, spiral, chevron, and circular arrangements. Comparison of tissues printed in a parallel alignment had more than double the contractile force of tissues printed with a perpendicular aOBB alignment. In the macrofilaments, aligned tissues exhibited higher contractile forces, sustained higher frequencies during electrical pacing, and demonstrated more synchronous action potentials compared to spheroid controls.160 While the aoBBs were not true cardioids, we can infer that similar usage of encapsulated cardioids printed using BATE would produce similar, if not better, functional outcomes. Similarly, Skylar-Scott et al.101 employed the SWIFT technique to print sacrificial channels into cardiac organ building blocks (OBBs) matrices containing iPSC-CMs and primary CFs. This approach produced high-density tissues (~240 million cells/mL) containing perfusable vascular channels, demonstrating how printed organoid matrices could be used to produce vascularized cardiac tissues.
For fabricating macroscale constructs, EBB has dominated the field, while LBB is preferred for high-resolution microarchitectures. Droplet-based bioprinting (DBB), however, struggles with large-tissue fabrication, making it less suited for cardioid bioprinting in its current state.161 Yet, advancements in biofabrication techniques and multi-modal printing strategies may soon enable high-fidelity, scalable printing of cardioids, expanding their clinical application. Volumetric bioprinting has recently emerged as a high-speed, high-resolution light-based modality that enables the creation of entire tissue constructs by projecting a series of optical patterns from multiple angles.162 While it has not been utilized with cardioids, volumetric bioprinting has already demonstrated geometrically complex, centimeter-scale liver and bone tissue using organoid-laden bioinks.163 Volumetric bioprinting and other hybrid approaches integrate developmental self-organization with engineering control, improving reproducibility, vascularization, and tissue scaling.
Whether printing cardioids or printing with cardioids, bioprinting enhances reproducibility and scalability, enabling automated, high-throughput production of cardiac tissues with uniform structure and composition.153 Commercially available bioinks have already demonstrated the ability to support functional cardiac tissue engineering in several studies. For example, Kupfer et al. 23 combined commercially sourced ColMA (Advanced BioMatrix) with GelMA to bioprint constructs exhibiting measurable contractile force, synchronous electrical activity, and perfusability. Wolfe et al.25 developed a patient-derived bioprinted patch for congenital heart disease modeling using GelMA Laminink-521, a commercially available bioink sold by CELLINK. Additionally, Jia et al.91 directly bioprinted perfusable vascular constructs using a blend bioink consisting of laboratory-prepared GelMA and PEGDA purchased from JenKem®. These examples are representative of the broader body of work demonstrating that commercially available bioinks can support functional cardiac tissue engineering. Additionally, these commercial bioinks can offer extensive quality control that may be difficult to achieve in a research environment. Although bioprinted cardioids are still in their infancy, improving scalability and reproducibility will be essential as organoids transition from research tools to clinical and commercial platforms.
5. Conclusion
Replicating the cellular, functional, and structural complexity of the native heart has been a longstanding challenge in cardiac tissue engineering. In this review, we examine how two concurrently evolving technologies—3D bioprinting and cardioid technology—independently address these challenges. Bioprinting offers scalability, spatial precision, and structural control, while cardioids provide multi-cellular diversity, developmental relevance, and physiological functionality.
While each approach has advanced the field, both face limitations that have restricted clinical and commercial translation, yet may be resolved through their convergence. Guiding or reinforcing cardioid self-organization through engineered spatial cues or using pre-formed cardioids as modular units within bioinks could generate more mature, vascularized, and functionally relevant cardiac tissues. Although cardiac bioprinting remains in its early stages, with challenges such as cellular immaturity, improper vascularization, and lack of scaling needing to be addressed, emerging proof-of-concept studies provide a strong foundation for future progress.
As both biofabrication techniques and cardiac stem cell differentiation protocols continue to advance, the integration of engineering and developmental biology will drive the next generation of in vitro cardiac models—supporting applications in developmental and disease modeling, drug screening, and ultimately cardiac repair and replacement.
1.Lindstrom M, DeCleene N, Dorsey H, et al. Global burden of cardiovascular diseases and risks collaboration, 1990–2021. J Am Coll Cardiol. 2022;80(25):2372-2425.
doi: 10.1016/j.jacc.2022.11.001
2.Cesare MD, Perel P, Taylor S, et al. The Heart of the World. Glob Heart. 2024;19(1):11.
doi: 10.5334/gh.1288
3.Tsao CW, Aday AW, Almarzooq ZI, et al. Heart disease and stroke statistics—2023 update: a report from the American Heart Association. Circulation. 2023;147(8):e93-e621.
doi: 10.1161/CIR.0000000000001123
4.Bergmann O, Bhardwaj RD, Bernard S, et al. Evidence for cardiomyocyte renewal in humans. Science. 2009;324(5923):98-102.
5.Heidenreich PA, Albert NM, Allen LA, et al. Forecasting the impact of heart failure in the United States. Circ Heart Fail. 2013;6(3):606-619.
doi: 10.1161/HHF.0b013e318291329a
6.Kazi DS, Elkind MSV, Deutsch A, et al. Forecasting the economic burden of cardiovascular disease and stroke in the United States through 2050: a presidential advisory from the American Heart Association. Circulation. 2024;150(4):e89-e101.
doi: 10.1161/CIR.0000000000001258
7.Thomas D, Choi S, Alamana C, Parker KK, Wu JC. Cellular and engineered organoids for cardiovascular models. Circ Res. 2022;130(12):1780-1802.
doi: 10.1161/CIRCRESAHA.122.320305
8.Cho S, Discher DE, Leong KW, Vunjak-Novakovic G, Wu JC. Challenges and opportunities for the next generation of cardiovascular tissue engineering. Nat Methods. 2022;19(9):1064-1071.
doi: 10.1038/s41592-022-01591-3
9.Wang Z, Wang L, Li T, et al. 3D bioprinting in cardiac tissue engineering. Theranostics. 2021;11(16):7948-7969.
doi: 10.7150/thno.61621
10.Sung K, Patel NR, Ashammakhi N, Nguyen KL. 3-Dimensional bioprinting of cardiovascular tissues: emerging technology. JACC: Basic Transl Sci. 2021;6(5):467-482.
doi: 10.1016/j.jacbts.2020.12.006
11.Zimmermann WH, Schneiderbanger K, Schubert P, et al. Tissue engineering of a differentiated cardiac muscle construct. Circ Res. 2002;90(2):223-230.
12.Wu F, He Q, Li F, Yang X. A review of protocols for engineering human cardiac organoids. Heliyon. 2023;9(9):e19938.
doi: 10.1016/j.heliyon.2023.e19938
13.Zhao D, Lei W, Hu S. Cardiac organoid—a promising perspective of preclinical model. Stem Cell Res Ther. 2021;12(1):272.
doi: 10.1186/s13287-021-02340-7
14.Zhu L, Liu K, Feng Q, Liao Y. Cardiac Organoids: A 3D technology for modeling heart development and disease. Stem Cell Rev Rep. 2022;18(8):2593-2605.
doi: 10.1007/s12015-022-10385-1
15.El Sabbagh A, Eleid MF, Al-Hijji M, et al. The various applications of 3D printing in cardiovascular diseases. Curr Cardiol Rep. 2018;20(6):47.
doi: 10.1007/s11886-018-0992-9
16.Hsieh PCH, Davis ME, Lisowski LK, Lee RT. Endothelial- cardiomyocyte interactions in cardiac development and repair. Annu Rev Physiol. 2006;68:51-66.
doi: 10.1146/annurev.physiol.68.040104.124629
17.Tian Y, Morrisey EE. Importance of myocyte-nonmyocyte interactions in cardiac development and disease. Circ Res. 2012;110(7):1023-1034.
doi: 10.1161/CIRCRESAHA.111.243899
18.Michel NA, Ljubojevic-Holzer S, Bugger H, Zirlik A. Cellular heterogeneity of the heart. Front Cardiovasc Med. 2022;9:868466.
19.Liu X, Huang J, Chen T, et al. Yamanaka factors critically regulate the developmental signaling network in mouse embryonic stem cells. Cell Res. 2008;18(12):1177-1189.
doi: 10.1038/cr.2008.309
20.Lyra-Leite DM, Gutiérrez-Gutiérrez Ó, Wang M, Zhou Y, Cyganek L, Burridge PW. A review of protocols for human iPSC culture, cardiac differentiation, subtype-specification, maturation, and direct reprogramming. STAR Protoc. 2022;3(3):101560.
doi: 10.1016/j.xpro.2022.101560
21.Lian X, Hsiao C, Wilson G, et al. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc Natl Acad Sci U S A. 2012;109(27):E1848-E1857.
22.Burridge PW, Matsa E, Shukla P, et al. Chemically defined and small molecule-based generation of human cardiomyocytes. Nat Methods. 2014;11(8):855-860.
doi: 10.1038/nmeth.2999
23.Kupfer ME, Lin WH, Ravikumar V, et al. In situ expansion, differentiation, and electromechanical coupling of human cardiac muscle in a 3D bioprinted, chambered organoid. Circ Res. 2020;127(2):207-224.
doi: 10.1161/CIRCRESAHA.119.316155
24.Esser TU, Anspach A, Muenzebrock KA, et al. Direct 3D-bioprinting of hiPSC-derived cardiomyocytes to generate functional cardiac tissues. Adv Mater. 2023; 35(52):2305911.
25.Wolfe JT, He W, Kim MS, et al. 3D-bioprinting of patient- derived cardiac tissue models for studying congenital heart disease. Front Cardiovasc Med. 2023;10:1162731.
doi: 10.3389/fcvm.2023.1162731
26.Köhne M, Behrens CS, Stüdemann T, et al. A potential future Fontan modification: preliminary in vitro data of a pressure-generating tube from engineered heart tissue. Eur J Cardiothorac Surg. 2022;62(2):ezac111.
27.MacQueen LA, Sheehy SP, Chantre CO, et al. A tissue- engineered scale model of the heart ventricle. Nat Biomed Eng. 2018;2(12):930-941.
doi: 10.1038/s41551-018-0271-5
28.Goldfracht I, Protze S, Shiti A, et al. Generating ring-shaped engineered heart tissues from ventricular and atrial human pluripotent stem cell-derived cardiomyocytes. Nat Commun. 2020;11(1):1-15.
doi: 10.1038/s41467-019-13868-x
29.Querdel E, Reinsch M, Castro L, et al. Human engineered heart tissue patches remuscularize the injured heart in a dose-dependent manner. Circulation. 2021;143(20): 1991-2006.
doi: 10.1161/CIRCULATIONAHA.120.047904
30.Mannhardt I, Breckwoldt K, Letuffe-Brenière D, et al. Human engineered heart tissue: analysis of contractile force. Stem Cell Rep. 2016;7(1):29-42.
doi: 10.1016/j.stemcr.2016.04.011
31.Jabbour RJ, Owen TJ, Pandey P, et al. In vivo grafting of large engineered heart tissue patches for cardiac repair. JCI Insight. 2021;6(15):e144068.
doi: 10.1172/jci.insight.144068
32.Lappi H, Kauppila M, Aalto-Setälä K, Mörö A. The 3D bioprinted human induced pluripotent stem cell-derived cardiac model: toward functional and patient-derived in vitro models for disease modeling and drug screening. Bioprinting. 2023;36:e00313.
doi: 10.1016/j.bprint.2023.e00313
33.Rehman S, Khan A, Rehman A. Physiology, coronary circulation. In: StatPearls. Treasure Island, FL: StatPearls Publishing; 2025.
http://www.ncbi.nlm.nih.gov/books/NBK482413/. Accessed January 10, 2025.
34.Maiullari F, Costantini M, Milan M, et al. A multi-cellular 3D bioprinting approach for vascularized heart tissue engineering based on HUVECs and iPSC-derived cardiomyocytes. Sci Rep. 2018;8(1):13532.
doi: 10.1038/s41598-018-31848-x
35.Zhang YS, Arneri A, Bersini S, et al. Bioprinting 3D microfibrous scaffolds for engineering endothelialized myocardium and heart-on-a-chip. Biomaterials. 2016;110:45-59.
doi: 10.1016/j.biomaterials.2016.09.003
36.Song HHG, Rumma RT, Ozaki CK, Edelman ER, Chen CS. Vascular tissue engineering: progress, challenges, and clinical promise. Cell Stem Cell. 2018;22(3): 40-354.
doi: 10.1016/j.stem.2018.02.009
37.Kennedy CC, Brown EE, Abutaleb NO, Truskey GA. Development and application of endothelial cells derived from pluripotent stem cells in microphysiological systems models. Front Cardiovasc Med. 2021;8:625016.
38.Gu M. Efficient differentiation of human pluripotent stem cells to endothelial cells. Curr Protoc Hum Genet. 2018;98(1):e64.
doi: 10.1002/cphg.64
39.Hamad S, Derichsweiler D, Gaspar JA, et al. High-efficient serum-free differentiation of endothelial cells from human iPS cells. Stem Cell Res Ther. 2022;13(1):251.
doi: 10.1186/s13287-022-02924-x
40.Williams IM, Wu JC. Generation of endothelial cells from human pluripotent stem cells. Arterioscler Thromb Vasc Biol. 2019;39(7):1317-1329.
doi: 10.1161/ATVBAHA.119.312265
41.Ayoubi S, Sheikh SP, Eskildsen TV. Human induced pluripotent stem cell-derived vascular smooth muscle cells: differentiation and therapeutic potential. Cardiovasc Res. 2017;113(11):1282-1293.
doi: 10.1093/cvr/cvx125
42.Souders CA, Bowers SLK, Baudino TA. Cardiac fibroblast: the renaissance cell. Circ Res. 2009;105(12):1164-1176.
doi: 10.1161/CIRCRESAHA.109.209809
43.Li RA, Keung W, Cashman TJ, et al. Bioengineering an electro-mechanically functional miniature ventricular heart chamber from human pluripotent stem cells. Biomaterials. 2018;163:116-127.
doi: 10.1016/j.biomaterials.2018.02.024
44.Tsuruyama S, Matsuura K, Sakaguchi K, Shimizu T. Pulsatile tubular cardiac tissues fabricated by wrapping human iPS cells-derived cardiomyocyte sheets. Regen Ther. 2019;11:297-305.
doi: 10.1016/j.reth.2019.09.001
45.Kawai Y, Tohyama S, Arai K, et al. Scaffold-free tubular engineered heart tissue from human induced pluripotent stem cells using bio-3D printing technology in vivo. Front Cardiovasc Med. 2022;8:806215.
46.Whitehead AJ, Hocker JD, Ren B, Engler AJ. Improved epicardial cardiac fibroblast generation from iPSCs. J Mol Cell Cardiol. 2022;164:58-68.
doi: 10.1016/j.yjmcc.2021.11.011
47.Zhang J, Tao R, Campbell KF, et al. Functional cardiac fibroblasts derived from human pluripotent stem cells via second heart field progenitors. Nat Commun. 2019;10(1):2238.
doi: 10.1038/s41467-019-09831-5
48.Bao X, Lian X, Qian T, Bhute VJ, Han T, Palecek SP. Directed differentiation and long-term maintenance of epicardial cells from human pluripotent stem cells under fully defined conditions. Nat Protoc. 2017;12(9):1890-1900.
49.Lee A, Hudson AR, Shiwarski DJ, et al. 3D bioprinting of collagen to rebuild components of the human heart. Science. 2019;365(6452):482-487.
50.Bliley JM, Vermeer MCSC, Duffy RM, et al. Dynamic loading of human engineered heart tissue enhances contractile function and drives a desmosome-linked disease phenotype. Sci Transl Med. 2021;13(603):eabd1817.
doi: 10.1126/scitranslmed.abd1817
51.Miller KL, Xiang Y, Yu C, et al. Rapid 3D BioPrinting of a human iPSC-derived cardiac micro-tissue for high-throughput drug testing. Organs Chip. 2021;3:100007.
doi: 10.1016/j.ooc.2021.100007
52.Fullenkamp DE, Maeng WY, Oh S, et al. Simultaneous electromechanical monitoring in engineered heart tissues using a mesoscale framework. Sci Adv.2024;10(37):eado7089.
53.Finkel S, Sweet S, Locke T, et al. FRESHTM 3D bioprinted cardiac tissue, a bioengineered platform for in vitro pharmacology. APL Bioeng. 2023;7(4):046113.
doi: 10.1063/5.0163363
54.Noor N, Shapira A, Edri R, Gal I, Wertheim L, Dvir T. 3D printing of personalized thick and perfusable cardiac patches and hearts. Adv Sci (Weinh). 2019;6(11):1900344.
55.Lock RI, Graney PL, Tavakol DN, et al. Macrophages enhance contractile force in iPSC-derived human engineered cardiac tissue. Cell Rep. 2024;43(6):114302.
doi: 10.1016/j.celrep.2024.114302
56.Szepes M, Melchert A, Dahlmann J, et al. Dual function of IPSC-derived pericyte-like cells in vascularization and fibrosis-related cardiac tissue remodeling in vitro. Int J Mol Sci. 2020;21(23):8947.
doi: 10.3390/ijms21238947
57.Karvinen J, Kellomäki M. Design aspects and characterization of hydrogel-based bioinks for extrusion-based bioprinting. Bioprinting. 2023;32:e00274.
doi: 10.1016/j.bprint.2023.e00274
58.Kabirian F, Mozafari M. Decellularized ECM-derived bioinks: prospects for the future. Methods. 2020;171:108-118.
doi: 10.1016/j.ymeth.2019.04.019
59.Gillispie G, Prim P, Copus J, et al. Assessment methodologies for extrusion-based bioink printability. Biofabrication. 2020;12(2):022003.
60.Blaeser A, Duarte Campos DF, Puster U, Richtering W, Stevens MM, Fischer H. Controlling shear stress in 3D bioprinting is a key factor to balance printing resolution and stem cell integrity. Adv Healthc Mater. 2016;5(3):326-333.
61.Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126(4):677-689.
doi: 10.1016/j.cell.2006.06.044
62.Caliari SR, Burdick JA. A practical guide to hydrogels for cell culture. Nat Methods. 2016;13(5):405-414.
doi: 10.1038/nmeth.3839
63.Lee KY, Mooney DJ. Alginate: properties and biomedical applications. Prog Polym Sci. 2012;37(1):106-126.
doi: 10.1016/j.progpolymsci.2011.06.003
64.Li Z, Ruan C, Niu X. Collagen-based bioinks for regenerative medicine: Fabrication, application and prospective. Med Nov Technol Devices. 2023;17:100211.
doi: 10.1016/j.medntd.2023.100211
65.Osidak EO, Kozhukhov VI, Osidak MS, Domogatsky SP. Collagen as bioink for bioprinting: a comprehensive review. Int J Bioprint. 2020;6(3):270.
66.Shpichka A, Osipova D, Efremov Y, et al. Fibrin-based bioinks: new tricks from an old dog. IJB. 2024;6(3):269. doi: 10.18063/ijb.v6i3.269
67.Kumar SA, Alonzo M, Allen SC, et al. A visible light-cross-linkable, fibrin–gelatin-based bioprinted construct with human cardiomyocytes and fibroblasts. ACS Biomater Sci Eng. 2019;5(9):4551-4563.
doi: 10.1021/acsbiomaterials.9b00505
68.Zhang D, Shadrin IY, Lam J, Xian HQ, Snodgrass HR, Bursac N. Tissue-engineered cardiac patch for advanced functional maturation of human ESC-derived cardiomyocytes. Biomaterials. 2013;34(23):5813-5820.
doi: 10.1016/j.biomaterials.2013.04.026
69.de Melo BAG, Jodat YA, Cruz EM, Benincasa JC, Shin SR, Porcionatto MA. Strategies to use fibrinogen as bioink for 3D bioprinting fibrin-based soft and hard tissues. Acta Biomater. 2020;117:60-76.
doi: 10.1016/j.actbio.2020.09.024
70.Bejleri D, Davis ME. Decellularized extracellular matrix materials for cardiac repair and regeneration. Adv Healthc Mater. 2019;8(5):1801217.
71.Jain P, Rauer SB, Möller M, Singh S. Mimicking the natural basement membrane for advanced tissue engineering. Biomacromolecules. 2022;23(8):3081-3103.
doi: 10.1021/acs.biomac.2c00402
72.Zhao KY, Du YX, Cao HM, Su LY, Su XL, Li X. The biological macromolecules constructed Matrigel for cultured organoids in biomedical and tissue engineering. Colloids Surf B Biointerfaces. 2025;247:114435.
doi: 10.1016/j.colsurfb.2024.114435
73.Kleinman HK, McGarvey ML, Liotta LA, Robey PG, Tryggvason K, Martin GR. Isolation and characterization of type IV procollagen, laminin, and heparan sulfate proteoglycan from the EHS sarcoma. Biochemistry. 1982;21(24):6188-6193.
doi: 10.1021/bi00267a025
74.Yue K, Santiago GT de, Alvarez MM, Tamayol A, Annabi N, Khademhosseini A. Synthesis, properties, and biomedical applications of gelatin methacryloyl (GelMA) hydrogels. Biomaterials. 2015;73:254-271.
doi: 10.1016/j.biomaterials.2015.08.045
75.Khoeini R, Nosrati H, Akbarzadeh A, et al. Natural and synthetic bioinks for 3D bioprinting. Adv NanoBiomed Res. 2021;1(8):2000097.
76.Wang X. Advanced polymers for three-dimensional (3D) organ bioprinting. Micromachines. 2019;10(12):814.
doi: 10.3390/mi10120814
77.Hockaday LA, Kang KH, Colangelo NW, et al. Rapid 3D printing of anatomically accurate and mechanically heterogeneous aortic valve hydrogel scaffolds. Biofabrication. 2012;4(3):035005.
doi: 10.1088/1758-5082/4/3/035005
78.Shin YJ, Shafranek RT, Tsui JH, Walcott J, Nelson A, Kim DH. 3D bioprinting of mechanically tuned bioinks derived from cardiac decellularized extracellular matrix. Acta Biomater. 2021;119:75-88.
doi: 10.1016/j.actbio.2020.11.006
79.Hersel U, Dahmen C, Kessler H. RGD modified polymers: biomaterials for stimulated cell adhesion and beyond. Biomaterials. 2003;24(24):4385-4415.
doi: 10.1016/s0142-9612(03)00343-0
80.Zhu J. Bioactive modification of poly(ethylene glycol) hydrogels for tissue engineering. Biomaterials. 2010;31(17):4639-4656.
doi: 10.1016/j.biomaterials.2010.02.044
81.Nourse MB, Halpin DE, Scatena M, et al. VEGF induces differentiation of functional endothelium from human embryonic stem cells. Arterioscler Thromb Vasc Biol. 2010;30(1):80-89.
doi: 10.1161/ATVBAHA.109.194233
82.Seliktar D, Zisch AH, Lutolf MP, Wrana JL, Hubbell JA. MMP-2 sensitive, VEGF-bearing bioactive hydrogels for promotion of vascular healing. J Biomed Mater Res A. 2004;68(4):704-716.
doi: 10.1002/jbm.a.20091
83.Balatskyi VV, Sowka A, Dobrzyn P, Piven OO. WNT/β- catenin pathway is a key regulator of cardiac function and energetic metabolism. Acta Physiol. 2023;237(3):e13912.
doi: 10.1111/apha.13912
84.Lee YB, Polio S, Lee W, et al. Bio-printing of collagen and VEGF-releasing fibrin gel scaffolds for neural stem cell culture. Exp Neurol. 2010;223(2):645-652.
doi: 10.1016/j.expneurol.2010.02.014
85.Poldervaart MT, Gremmels H, van Deventer K, et al. Prolonged presence of VEGF promotes vascularization in 3D bioprinted scaffolds with defined architecture. J Control Release. 2014;184:58-66.
doi: 10.1016/j.jconrel.2014.04.007
86.Mann BK, Schmedlen RH, West JL. Tethered- TGF-β increases extracellular matrix production of vascular smooth muscle cells. Biomaterials. 2001; 22(5):439-444.
doi: 10.1016/S0142-9612(00)00196-4
87.DeLong SA, Moon JJ, West JL. Covalently immobilized gradients of bFGF on hydrogel scaffolds for directed cell migration. Biomaterials. 2005;26(16):3227-3234.
doi: 10.1016/j.biomaterials.2004.09.021
88.Bliley J, Tashman J, Stang M, et al. FRESH 3D bioprinting a contractile heart tube using human stem cell-derived cardiomyocytes. Biofabrication. 2022;14(2):10.1088/1758- 5090/ac58be.
89.Ren J, Han P, Ma X, et al. Canonical Wnt5b signaling directs outlying Nkx2.5+ mesoderm into pacemaker cardiomyocytes. Dev Cell. 2019;50(6):729-743.e5.
doi: 10.1016/j.devcel.2019.07.014
90.Liu J, He J, Liu J, et al. Rapid 3D bioprinting of in vitro cardiac tissue models using human embryonic stem cell-derived cardiomyocytes. Bioprinting. 2019;13:e00040.
doi: 10.1016/j.bprint.2019.e00040
91.Jia W, Gungor-Ozkerim PS, Zhang YS, et al. Direct 3D bioprinting of perfusable vascular constructs using a blend bioink. Biomaterials. 2016;106:58-68.
doi: 10.1016/j.biomaterials.2016.07.038
92.Pati F, Jang J, Lee JW, Cho DW. Chapter 7—extrusion bioprinting. In: Atala A, Yoo JJ, eds. Essentials of 3D Biofabrication and Translation. Academic Press; 2015:123-152.
doi: 10.1016/B978-0-12-800972-7.00007-4
93.Li H, Dai J, Wang Z, et al. Digital light processing (DLP)- based (bio)printing strategies for tissue modeling and regeneration. Aggregate. 2023;4(2):e270.
doi: 10.1002/agt2.270
94.Hosseinabadi HG, Dogan E, Miri AK, Ionov L. Digital light processing bioprinting advances for micro-tissue models. ACS Biomater Sci Eng. 2022;8(4):1381-1395.
doi: 10.1021/acsbiomaterials.1c01509
95.Gudapati H, Dey M, Ozbolat I. A comprehensive review on droplet-based bioprinting: past, present and future. Biomaterials. 2016;102:20-42.
doi: 10.1016/j.biomaterials.2016.06.012
96.Shin S, Kwon Y, Hwang C, et al. Visible-light-driven rapid 3d printing of photoresponsive resins for optically clear multifunctional 3D objects. Adv Mater. 2024;36(19):2311917.
97.Salem T, Frankman Z, Churko JM. Tissue engineering techniques for induced pluripotent stem cell derived three-dimensional cardiac constructs. Tissue Eng Part B Rev. 2022;28(4):891-911.
doi: 10.1089/ten.teb.2021.0088
98.You S, Xiang Y, Hwang HH, et al. High cell density and high-resolution 3D bioprinting for fabricating vascularized tissues. Sci Adv. 2023;9(8):eade7923.
99.Hinton TJ, Jallerat Q, Palchesko RN, et al. Three-dimensional printing of complex biological structures by freeform reversible embedding of suspended hydrogels. Sci Adv. 2015;1(9):e1500758.
100.Mirdamadi E, Tashman JW, Shiwarski DJ, Palchesko RN, Feinberg AW. FRESH 3D bioprinting a full-size model of the human heart. ACS Biomater Sci Eng. 2020;6(11): 6453-6459.
doi: 10.1021/acsbiomaterials.0c01133
101.Skylar-Scott MA, Uzel SGM, Nam LL, et al. Biomanufacturing of organ-specific tissues with high cellular density and embedded vascular channels. Sci Adv. 2019;5(9):eaaw2459.
102.Bhusal A, Dogan E, Nguyen HA, et al. Multi-material digital light processing bioprinting of hydrogel-based microfluidic chips. Biofabrication. 2021;14(1):014103.
103.Liu J, Miller K, Ma X, et al. Direct 3D bioprinting of cardiac micro-tissues mimicking native myocardium. Biomaterials. 2020;256:120204.
doi: 10.1016/j.biomaterials.2020.120204
104.Yu C, Ma X, Zhu W, et al. Scanningless and continuous 3D bioprinting of human tissues with decellularized extracellular matrix. Biomaterials. 2019;194:1-13.
doi: 10.1016/j.biomaterials.2018.12.009
105.Zhu W, Qu X, Zhu J, et al. Direct 3D bioprinting of prevascularized tissue constructs with complex microarchitecture. Biomaterials. 2017;124:106-115.
doi: 10.1016/j.biomaterials.2017.01.042
106.Izadifar M, Babyn P, Kelly ME, Chapman D, Chen X. Bioprinting pattern-dependent electrical/mechanical behavior of cardiac alginate implants: characterization and ex vivo phase-contrast microtomography assessment. Tissue Eng Part C Methods. 2017;23(9):548-564.
doi: 10.1089/ten.tec.2017.0222
107.Kotadia I, Whitaker J, Roney C, et al. Anisotropic cardiac conduction. Arrhythm Electrophysiol Rev. 2020;9(4):202-210.
doi: 10.15420/aer.2020.04
108.Bera AK, Rizvi MS, KN V, Pati F. Engineering anisotropic tissue analogues: harnessing synergistic potential of extrusion-based bioprinting and extracellular matrix-based bioink. Biofabrication. 2024;17(1):015003.
109.Dwyer KD, Coulombe KLK. Cardiac mechanostructure: using mechanics and anisotropy as inspiration for developing epicardial therapies in treating myocardial infarction. Bioact Maters. 2021;6(7):2198-2220.
doi: 10.1016/j.bioactmat.2020.12.015
110.Chikae S, Kubota A, Nakamura H, et al. Three-dimensional bioprinting human cardiac tissue chips of using a painting needle method. Biotechnol Bioeng. 2019;116(11):3136-3142.
doi: 10.1002/bit.27126
111.Xu T, Baicu C, Aho M, Zile M, Boland T. Fabrication and characterization of bio-engineered cardiac pseudo tissues. Biofabrication. 2009;1(3):035001.
doi: 10.1088/1758-5082/1/3/035001
112.El Khoury R, Nagiah N, Mudloff JA, Thakur V, Chattopadhyay M, Joddar B. 3D bioprinted spheroidal droplets for engineering the heterocellular coupling between cardiomyocytes and cardiac fibroblasts. Cyborg Bionic Syst. 2021;2021:9864212.
113.Datta P, Ayan B, Ozbolat IT. Bioprinting for vascular and vascularized tissue biofabrication. Acta Biomater. 2017;51:1-20.
doi: 10.1016/j.actbio.2017.01.035
114.Elkhoury K, Zuazola J, Vijayavenkataraman S. Bioprinting the future using light: a review on photocrosslinking reactions, photoreactive groups, and photoinitiators. SLAS Technol. 2023;28(3):142-151.
doi: 10.1016/j.slast.2023.02.003
115.GhavamiNejad A, Ashammakhi N, Wu XY, Khademhosseini A. Crosslinking strategies for three-dimensional bioprinting of polymeric hydrogels. Small. 2020;16(35):e2002931.
116.Lee JE, Heo SW, Kim CH, Park SJ, Park SH, Kim TH. In-situ ionic crosslinking of 3D bioprinted cell-hydrogel constructs for mechanical reinforcement and improved cell growth. Biomater Adv. 2023;147:213322.
doi: 10.1016/j.bioadv.2023.213322
117.Chatterjee S, Hui PCL, Kan CW, Wang W. Dual-responsive (pH/temperature) Pluronic F-127 hydrogel drug delivery system for textile-based transdermal therapy. Sci Rep. 2019;9(1):11658.
doi: 10.1038/s41598-019-48254-6
118.Sarker M, Izadifar M, Schreyer D, Chen X. Influence of ionic crosslinkers (Ca2+/Ba2+/Zn2+) on the mechanical and biological properties of 3D bioplotted hydrogel scaffolds. J Biomater Sci Polym Ed. 2018;29(10):1126-1154.
doi: 10.1080/09205063.2018.1433420
119.Kim MH, Lee YW, Jung WK, Oh J, Nam SY. Enhanced rheological behaviors of alginate hydrogels with carrageenan for extrusion-based bioprinting. J Mech Behav Biomed Mater. 2019;98:187-194.
doi: 10.1016/j.jmbbm.2019.06.014
120.Gu Y, Zhang L, Du X, et al. Reversible physical crosslinking strategy with optimal temperature for 3D bioprinting of human chondrocyte-laden gelatin methacryloyl bioink. J Biomater Appl. 2018;33(5):609-618.
121.Bahney CS, Lujan TJ, Hsu CW, Bottlang M, West JL, Johnstone B. Visible light photoinitiation of mesenchymal stem cell-laden bioresponsive hydrogels. Eur Cell Mater. 2011;22:43-55; discussion 55.
doi: 10.22203/ecm.v022a04
122.Basara G, Ozcebe SG, Ellis BW, Zorlutuna P. Tunable Human myocardium derived decellularized extracellular matrix for 3D bioprinting and cardiac tissue engineering. Gels. 2021;7(2):70.
doi: 10.3390/gels7020070
123.Budharaju H, Sundaramurthi D, Sethuraman S. Efficient dual crosslinking of protein–in–polysaccharide bioink for biofabrication of cardiac tissue constructs. Biomater Adv. 2023;152:213486.
doi: 10.1016/j.bioadv.2023.213486
124.Karbassi E, Fenix A, Marchiano S, et al. Cardiomyocyte maturation: advances in knowledge and implications for regenerative medicine. Nat Rev Cardiol. 2020; 17(6):341-359.
doi: 10.1038/s41569-019-0331-x
125.Wu P, Deng G, Sai X, Guo H, Huang H, Zhu P. Maturation strategies and limitations of induced pluripotent stem cell-derived cardiomyocytes. Biosci Rep. 2021;41(6):BSR20200833.
doi: 10.1042/BSR20200833
126.Guo Y, Pu W. Cardiomyocyte maturation: new phase in development. Circ Res. 2020;126(8):1086-1106.
doi: 10.1161/CIRCRESAHA.119.315862
127.Tulloch NL, Muskheli V, Razumova MV, et al. Growth of engineered human myocardium with mechanical loading and vascular coculture. Circ Res. 2011;109(1):47-59.
doi: 10.1161/CIRCRESAHA.110.237206
128.Zhao Y, Rafatian N, Feric NT, et al. A platform for generation of chamber specific cardiac tissues and disease modelling. Cell. 2019;176(4):913-927.e18.
doi: 10.1016/j.cell.2018.11.042
129.Ronaldson-Bouchard K, Ma SP, Yeager K, et al. Advanced maturation of human cardiac tissue grown from pluripotent stem cells. Nature. 2018;556(7700):239-243.
doi: 10.1038/s41586-018-0016-3
130.Godier-Furnémont AFG, Tiburcy M, Wagner E, et al. Physiologic force-frequency response in engineered heart muscle by electromechanical stimulation. Biomaterials. 2015;60:82-91.
doi: 10.1016/j.biomaterials.2015.03.055
131.Scuderi GJ, Butcher J. Naturally engineered maturation of cardiomyocytes. Front Cell Dev Biol. 2017;5:50.
132.Bliley JM, Stang MA, Behre A, Feinberg AW. Advances in 3D bioprinted cardiac tissue using stem cell-derived cardiomyocytes. Stem Cells Transl Med. 2024;13(5):425-435.
133.Paez-Mayorga J, Hernández-Vargas G, Ruiz-Esparza GU, et al. Bioreactors for cardiac tissue engineering. Adv Healthc Mater. 2019;8(7):1701504.
134.Kim H, Kamm RD, Vunjak-Novakovic G, Wu JC. Progress in multicellular human cardiac organoids for clinical applications. Cell Stem Cell. 2022;29(4):503-514.
doi: 10.1016/j.stem.2022.03.012
135.Drakhlis L, Biswanath S, Farr CM, et al. Human heart-forming organoids recapitulate early heart and foregut development. Nat Biotechnol. 2021;39(6):737-746.
doi: 10.1038/s41587-021-00815-9
136.Lewis-Israeli YR, Wasserman AH, Gabalski MA, et al. Self-assembling human heart organoids for the modeling of cardiac development and congenital heart disease. Nat Commun. 2021;12(1):5142.
doi: 10.1038/s41467-021-25329-5
137.Rossi G, Broguiere N, Miyamoto M, et al. Capturing cardiogenesis in gastruloids. Cell Stem Cell. 2021;28(2):230-240.e6.
doi: 10.1016/j.stem.2020.10.013
138.Hofbauer P, Jahnel SM, Papai N, et al. Cardioids reveal self-organizing principles of human cardiogenesis. Cell. 2021;184(12):3299-3317.e22.
doi: 10.1016/j.cell.2021.04.034
139.Silva AC, Matthys OB, Joy DA, et al. Co-emergence of cardiac and gut tissues promotes cardiomyocyte maturation within human iPSC-derived organoids. Cell Stem Cell. 2021;28(12):2137-2152.e6.
doi: 10.1016/j.stem.2021.11.007
140.Branco MA, Dias TP, Cabral JMS, Pinto-do-Ó P, Diogo MM. Human multilineage pro-epicardium/foregut organoids support the development of an epicardium/myocardium organoid. Nat Commun. 2022;13(1):6981.
doi: 10.1038/s41467-022-34730-7
141.Olmsted ZT, Paluh JL. A combined human gastruloid model of cardiogenesis and neurogenesis. iScience. 2022;25(6):104486.
doi: 10.1016/j.isci.2022.104486
142.Meier AB, Zawada D, De Angelis MT, et al. Epicardioid single-cell genomics uncovers principles of human epicardium biology in heart development and disease. Nat Biotechnol. 2023;41(12):1787-1800.
doi: 10.1038/s41587-023-01718-7
143.Volmert B, Kiselev A, Juhong A, et al. A patterned human primitive heart organoid model generated by pluripotent stem cell self-organization. Nat Commun. 2023; 14(1):8245.
doi: 10.1038/s41467-023-43999-1
144.Mohr E, Thum T, Bär C. Accelerating cardiovascular research: recent advances in translational 2D and 3D heart models. Eur J Heart Fail. 2022;24(10):1778-1791.
doi: 10.1002/ejhf.2631
145.Kehat I, Kenyagin-Karsenti D, Snir M, et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest. 2001;108(3):407-414.
doi: 10.1172/JCI12131
146.Burridge PW, Anderson D, Priddle H, et al. Improved human embryonic stem cell embryoid body homogeneity and cardiomyocyte differentiation from a novel v-96 plate aggregation system highlights interline variability. Stem Cells. 2007;25(4):929-938.
doi: 10.1634/stemcells.2006-0598
147.Voges HK, Mills RJ, Elliott DA, Parton RG, Porrello ER, Hudson JE. Development of a human cardiac organoid injury model reveals innate regenerative potential. Development. 2017;144(6):1118-1127.
doi: 10.1242/dev.143966
148.Lemon LS, Bodnar LM, Garrard W, et al. Ondansetron use in the first trimester of pregnancy and the risk of neonatal ventricular septal defect. Int J Epidemiol. 2020;49(2): 648-656.
doi: 10.1093/ije/dyz255
149.Richards DJ, Li Y, Kerr CM, et al. Human cardiac organoids for the modelling of myocardial infarction and drug cardiotoxicity. Nat Biomed Eng. 2020;4(4):446-462.
doi: 10.1038/s41551-020-0539-4
150.Octavia Y, Tocchetti CG, Gabrielson KL, Janssens S, Crijns HJ, Moens AL. Doxorubicin-induced cardiomyopathy: from molecular mechanisms to therapeutic strategies. J Mol Cell Cardiol. 2012;52(6):1213-1225.
doi: 10.1016/j.yjmcc.2012.03.006
151.Brassard JA, Nikolaev M, Hübscher T, Hofer M, Lutolf MP. Recapitulating macro-scale tissue self-organization through organoid bioprinting. Nat Mater. 2021;20(1):22-29.
doi: 10.1038/s41563-020-00803-5
152.Skylar-Scott MA, Huang JY, Lu A, et al. Orthogonally induced differentiation of stem cells for the programmatic patterning of vascularized organoids and bioprinted tissues. Nat Biomed Eng. 2022;6(4):449-462.
doi: 10.1038/s41551-022-00856-8
153.Lawlor KT, Vanslambrouck JM, Higgins JW, et al. Cellular extrusion bioprinting improves kidney organoid reproducibility and conformation. Nat Mater. 2021;20(2):260-271.
doi: 10.1038/s41563-020-00853-9
154.Yang H, Sun L, Pang Y, et al. Three-dimensional bioprinted hepatorganoids prolong survival of mice with liver failure. Gut. 2021;70(3):567-574.
doi: 10.1136/gutjnl-2019-319960
155.Choi Y mi, Lee H, Ann M, Song M, Rheey J, Jang J. 3D bioprinted vascularized lung cancer organoid models with underlying disease capable of more precise drug evaluation. Biofabrication. 2023;15(3):034104.
156.Wang D, Guo Y, Zhu J, et al. Hyaluronic acid methacrylate/ pancreatic extracellular matrix as a potential 3D printing bioink for constructing islet organoids. Acta Biomater. 2023;165:86-101.
doi: 10.1016/j.actbio.2022.06.036
157.He C, Yan J, Fu Y, Guo J, Shi Y, Guo J. Organoid bioprinting strategy and application in biomedicine: a review. IJB. 2023;9(6):0112.
doi: 10.36922/ijb.0112
158.Liu F, Liu C, Chen Q, et al. Progress in organ 3D bioprinting. Int J Bioprint. 2018;4(1):128.
159.Romanazzo S, Nemec S, Roohani I. iPSC bioprinting: where are we at? Materials (Basel). 2019;12(15):2453.
doi: 10.3390/ma12152453
160.Ahrens JH, Uzel SGM, Skylar‐Scott M, et al. Programming cellular alignment in engineered cardiac tissue via bioprinting anisotropic organ building blocks. Adv Mater. 2022;34(26):2200217.
161.Ren Y, Yang X, Ma Z, et al. Developments and opportunities for 3D bioprinted organoids. Int J Bioprint. 2021;7(3):364.
162.Bernal PN, Delrot P, Loterie D, et al. Volumetric bioprinting of complex living-tissue constructs within seconds. Adv Mater. 2019;31(42):1904209.
163.Bernal PN, Bouwmeester M, Madrid-Wolff J, et al. Volumetric bioprinting of organoids and optically tuned hydrogels to build liver-like metabolic biofactories. Adv Mater. 2022;34(15):2110054.